| EGGE-WESER | Bd. 7 | Seite: 39-56 | Höxter 1995 |
MORPHOLOGISCHE DIFFERENZIERUNG
DER ORCHIS MASCULA
IN ABHÄNGIGKEIT VON DER VARIABILITÄT
STANDÖRTLICHER GEGEBENHEITEN
Christian Evertz und Lars Meckelnburg
Inhalt
- VORWORT
- KURZFASSUNG
- EINLEITUNG
- GEOGRAPHISCHE UND TOPOGRAPHISCHE LAGEBESCHREIBUNG DES UNTERSUCHUNGSGEBIETES
- GEOLOGISCH-GEOMORPHOLOGISCHE STRUKTUR DES UNTERSUCHUNGSGEBIETES
- KLIMATISCHE GEGEBENHEITEN DES UNTERSUCHUNGSGEBIETES
- BODENFORMATION DES UNTERSUCHUNGSGEBIETES
- ARBEITSWEISEN, ARBEITSVERFAHREN UND ZIELSETZUNG DER UNTERSUCHUNG
- BESCHREIBUNG DER FORMENVIELFALT BEIM STATTLICHEN KNABENKRAUT (ORCHIS MASCULA)
- INTERPRETATION DER AUSWERTUNG
- TAXONOMIE UND SYSTEMATIK
- MAßNAHMEN ZUR ERHALTUNG DES BESTANDES
- LITERATURNACHWEIS
Vorwort
Bedingt durch die Tatsache, daß das Weserbergland für seinen Orchideenreichtum bekannt ist und in der Nähe unserer Heimatstadt große Orchideenvorkommen mit einer enormen Formenvielfalt auf engem Raum zu verzeichnen sind, stellten wir uns im Rahmen unserer Arbeit die Frage, ob eine modifikatorisch bedingte morphologische Differenzierung von Orchis mascula stattfinden kann. In der gängigen Fachliteratur ist bei anderen Orchideenarten die Rede von Standortmodifikationen, nicht aber bei Orchis mascula.
Unsere Untersuchungen beschränken sich auf ein 2 ha großes Areal, auf dem 500 Orchideen zu verzeichnen waren. Da dieses Gebiet noch nicht unter Naturschutz steht, kommen wir dem Wunsch des Betreuers dieses Areals nach, keine genaue Lagebeschreibung anzugeben.
Wir möchten an dieser Stelle recht herzlich unserem Betreuungslehrer OStR Hans Barth danken, der uns mit seinen fachkundigen Ratschlägen zur Seite stand und bereitwillig seine kostbare Zeit opferte.
Kurzfassung
Orchideen sind mit ihrem Alter von 15 Millionen Jahren bekanntlich eine der jüngsten Pflanzenfamilien, was im Gegensatz zu anderen Pflanzenfamilien, die bereits auf 100 Millionen Jahre Geschichte zurückblicken können, zu einer wenig differenzierten Abgrenzung von nahverwandten Arten führt. Aufgrund dieser Tatsache gibt es eine Fülle von Artengruppen innerhalb der Orchidales, die oftmals durch fließende Übergänge eine phylogenetische Unterscheidung äußerst schwierig gestalten lassen. Ein hervorragendes Beispiel für die überaus hohe Variabilität stellt Orchis mascula dar, die auf einer rund 2 ha großen Fläche im südöstlichen Weserbergland in einer bemerkenswert großen Population vorkommt. Ziel der Untersuchung ist es, die enorme Formenvielfalt bei Orchis mascula, da sie in den gängigen Exkursionsfloren nirgendwo erwähnt wird, zu erfassen und darzustellen. Man hat das Untersuchungsgebiet in drei Zonen geteilt, die durch ihre Vegetation innerhalb jeder Zone bestimmte abiotische Faktoren aufweisen. Orchis mascula zeigt auf der Untersuchungsfläche von rund zwei Hektar eine enorme Variabilität in der Gestaltung des Cormus (vgl. Abb. l/ Abb. 4). Die Merkmalsdifferenzierung reicht von kurzsprossigen zu langsprossigen Exemplaren. Dabei treten Unterschiede von 43 cm auf. Vor allem die Unterschiede in der Farbintensität der Einzelblüten ist hierbei auffällig. Das Farbenspektrum reicht von blaßviolett, in Einzelfällen von weißlichen Blüten, über violette Formen zu tiefvioletten dunklen Varianten. Betrachtet man den Gesamthabitus der einzelnen Orchis mascula-Formen, so
lassen sich immer wieder in Erscheinung tretende Merkmalspaarungen oder Kombinationen spezifisch morphologischer Strukturen nachvollziehen. Die Merkmalsdifferenzierung erfolgt kontinuierlich. Durch die abiotischen Faktoren findet eine Anpassung der Orchis mascula statt, was z. B. die Intensität des Lichtes belegt: Es tritt die charakteristische Erscheinung der Vergeilung auf. Auch wenn bei den Monokotyledonen wie bei Orchis mascula weniger die Länge der Sproßachse als vielmehr die Länge der Blätter verändert wird, so weist das Längenwachstum der Orchis mascula deutlich auf einen Lichtmangel hin. Es findet also durch die Veränderung der abiotischen Faktoren, bezogen auf den Standort der Orchideen, eine Metamorphose von Blättern und Sproß statt. Selbst unter der Berücksichtigung, daß genetische Untersuchungen nicht angestellt wurden und das Untersuchungsgebiet als kleinräumig anzusehen ist, kann man insgesamt dennoch von einer ökologischen Differenzierung des Artbereichs durch eine Öko-Cline-Bildung sprechen. Da davon auszugehen ist, daß die Familie der Orchidaceen evolutiv gesehen sehr jung ist und eine typische schrittweise Merkmalsverschiebung schneller zur Wirkung kommt, herrscht im Fall des Untersuchungsgebietes eine dynamische Selektion vor, die nur dann in eine stabilisierende übergehen kann, wenn die drei unterschiedlichen ökologischen Nischen über einen langen Zeitraum erhalten werden.
Die Orchis mascula-Population zeigt auf engstem Raum innerhalb des Untersuchungsgebietes eine Formenvielfalt, die zunächst auf eine Aufspaltung des Genpools schließen läßt. Doch die stufenweise Merkmalsdifferenzierung in Form einer Cline-Ausbildung belegt eindeutig, daß der Genfluß zwischen den einzelnen Teilpopulationen aufrechterhalten bleibt. Um jedoch endgültigen Aufschluß über die taxonomische Gliederung der formenreichen Art befriedigend lösen zu können, bedarf es daher verschiedener genetischer Untersuchungen, die im Rahmen dieser Untersuchung nicht zu leisten waren. Es handelt sich aber aller Wahrscheinlichkeit nach bei den untersuchten De-men von Orchis mascula um Standortmodifikationen. Da verschiedene Formen von Orchis mascula in einzelnen Herden gemeinsam auftreten, befindet sich die Orchis mascula-Population in der Aufspaltung unter dem Niveau der Lokalrassen.
Einleitung
Die Klärung der Frage, wie viele Orchideen es in Europa gibt, ist bei Systematikern, Geobotanikern und Phylogenetikern oft mit einem Glaubenskrieg verbunden. Je nachdem wie und was gezählt wird, sind die Unterschiede in der Artenzahl erheblich. Die dabei ausgewiesene Zahl reicht von 105 bis 246 Arten, die vielen Unterarten dabei nicht eingeschlossen. Gründe für diese große Spanne liegen zum einen im hohen Verwandtschaftsgrad und dem damit verbundenen hohen Ähnlichkeitsgrad zahlreicher Orchideen, zum anderen in den verschiedenen Methoden der Bestimmung begründet.
Orchideen sind mit ihrem Alter von 15 Millionen Jahren bekanntlich eine der jüngsten Blütenpflanzenfamilien, was im Gegensatz zu anderen Pflanzenfamilien, die bereits auf eine 100 Millionen Jahre alte Geschichte zurückblicken können, zu einer wenig differenzierten Abgrenzung von nah verwandten Arten führt. Aufgrund dieser Tatsache gibt es eine Fülle von Artengruppen innerhalb der Orchidaceae, die oftmals durch fließende Übergänge eine Unterscheidung äußerst schwierig gestalten lassen.
Ein hervorragendes Beispiel für die überaus hohe Variabilität stellt das Stattliche Knabenkraut (Orchis mascula) dar, das auf einer rund 2 ha großen Fläche im südöstlichen Weserbergland in einer bemerkenswert großen Population vorkommt. Ihr Formenreichtum, der bei einer oberflächlichen Betrachtung den Anschein erweckt, daß es sich hierbei um verschiedene Rassen, wenn nicht sogar um unterschiedliche Arten der Familie der Knabenkräuter (Orchidaceae) handelt, ist der Anlaß einer Untersuchung gewesen, die für die morphologische Abstufung (Abstufung in der äußeren Erscheinungsform) innerhalb des Formenkreises verantwortlich sind.
Geographische und topographische Lagebeschreibung des Untersuchungsgebietes
Das Untersuchungsgebiet liegt in der Nähe von Amelunxen (Stadt Beverungen, Kreis Höxter) im Wuchsgebiet des Weserberglandes. Die Kartierungsgebiete liegen in einem rund 2 ha großen Areal des ansonsten 100 ha großen Untersuchungsgebietes. Das gesamte Areal dieses Raums, welches eine durchschnittliche Höhe von rund 240 m ü.NN aufweist, ist heute im Besitz der Amelunxer Waldgenossenschaft und wird vom Staatlichen Forstamt Bad Driburg betreut.
Geologisch-geomorphologische Struktur des Untersuchungsgebietes
Die Untersuchungsfläche befindet sich auf einem Plateau des Scheelenberges, der sich mit seiner sargdeckelförmigen Struktur und seinen Schichtstufen in das System der Mittelgebirgsformation des Weserberglandes einordnet. Dieser Bergrücken gehört einer Hochfläche an, die durch fluviatile Erosion (Abtrag durch Wasser) aus dem Gebirgssystem gelöst wurde. Die im Norden und Süden in Ost-West-Richtung verlaufenden Talungen sowie die im Westen und Osten von Norden nach Süden eingesenkten Täler trennen somit in markanter Weise diese Fläche von den in ihrer Gesteinsformation sehr ähnlichen Oberflächenstrukturen ab. Im Bereich des Plateaus kam es vereinzelt durch Zerschneidungen zur Auflösung der flächenhaften Oberflächenform, so daß als Ergebnis trapezförmige Gebirgsrücken entstanden, die aber durch ihren plateauhaften Charakter immer noch deutlich auf eine ehemals homogene Hochfläche schließen lassen. Weniger innere als vielmehr äußere Kräfte sind es gewesen, die für die Oberflächenstruktur (Relief) verantwortlich waren. Für die tafelbergähnliche Gestalt sind jedoch nicht ausschließlich äußere Kräfte als Ursache anzuführen, sondern die Gesteinsformationen der Fläche selbst. Die in der Trias (vor 225-205 Mio. Jahren) horizontal zur Ablagerung gekommenen Sedimente des Buntsandsteins, Muschelkalks und Keupers verfestigten sich und bildeten Sedimentgesteine.
Entsprechend der Abfolge der jeweiligen geologischen Epochen tritt am Hangfußbereich der fluviatil angeschnittene obere Buntsandstein zu Tage, der weiter im mittleren Hangbereich in die mächtigen Formationen des unteren Muschelkalkes übergeht. Dieser wird wiederum von einem schmalen Band des Mittleren Muschelkalkes abgelöst. Der Oberhang setzt sich in Fortführung der chronologischen Folge der Trias aus Gesteinsschichten des Oberen Muschelkalkes zusammen, der im inneren Bereich, allerdings regional engbegrenzt, von Formationen des Unteren Keupers überlagert wird. Aufgrund der Hanglänge sowie der konkaven Hangstruktur wurden die Schichten des Keupers erodiert, so daß die Decken des Oberen Muschelkalkes in den Randbereichen der Hochfläche den Abschluß darstellen. Darüber hinaus rief der strenge Wechsel von harten und weichen Gesteinsschichten der einzelnen Formationen eine treppenartige Stufung der Hänge hervor, indem die weicheren Gesteine abgetragen wurden und somit die härteren Schichten die Hangstruktur bestimmen. Diese als Schichtstufen zu bezeichnende geomorphologische Erscheinungsform verdankt somit ihre Entstehung den widerstandsfähigen Schichten des Unteren und Oberen Muschelkalkes. Sie werden daher als Stufenbildner bezeichnet.
Diese spezifische Anordnung der Gesteinsabfolge finden wir ebenfalls im südlichen Hangbereich vor, in dem auch das Untersuchungsgebiet gelegen ist. Da sich die Untersuchungsfläche an der Hangoberkante befindet und sich in nordwestlicher Richtung auf das Plateau ausdehnt, wird ihr Untergrund von dem sehr widerstandsfähigen Oberen Muschelkalk bestimmt. Seine oberflächennah anstehenden Gesteinsschichten, die durch chemische Verwitterung bankig zerfallen sind, bedecken als Schuttmantel die darunterliegenden, ebenfalls zum Oberen Muschelkalk gehörenden Schichten.
Klimatische Gegebenheiten des Untersuchungsgebietes
Das auf die Fläche einwirkende Klima weist in seinen Grundstrukturen eine ozeanische Struktur auf, die aber, wie die kurzen Sommer und langen Winter belegen, eine leicht kontinentale Tönung erfährt. Durch die
fehlenden Barrieren im Osten dringen kontinentale Luftmassen aus Zentralrußland in das Weserbergland vor und bringen im Sommer Wärme und im Winter Kälte mit. Sie intensivieren somit die Extrema der auf die Region wirkenden Klimabedingungen. Das südliche Weserbergland wird deshalb zum Klimabereich Westliches Mitteldeutschland bzw. zum Klimabezirk "Oberes Bergland" gerechnet. Mit mittleren Niederschlägen von rund 750 mm/a ist die Region und damit auch die Umgebung der Untersuchungsfläche als niederschlagsarmer Bereich auszuweisen. Auffällig dabei ist, daß der März mit einer durchschnittlichen Niederschlagsmenge von nur 40 mm den trockensten, die Monate Juli und August dagegen die niederschlagsreichsten Monate darstellen. Die Auswirkungen der geringen Niederschlagsmengen werden durch die Tatsache verstärkt, daß die jährlich aktuelle Verdunstung zwischen 450 und 500 mm/a liegt. Auf die Untersuchungsfläche bezogen, die an einem südexponierten Hang gelegen ist, dürfte wohl der Oberwert von rund 500 mm/a angenommen werden, so daß die Differenz von 250 mm/a die Trockenanfälligkeit der Fläche deutlich unterstreicht. Die mittlere Lufttemperatur liegt im Januar bei 0°C und im Juli bei 16°C. Die Durchschnittstemperatur wird mit 8°C angegeben. Die mittlere Zahl der Frosttage im Jahr schwankt zwischen 80 und 100, wobei aufgrund der exponierten Lage auch hier der höhere Wert angenommen werden kann. Die Dauer der frostfreien Periode umfaßt einen Zeitraum von 149 bis 192 Tagen.
Aufgrund der klimatischen Gegebenheiten erscheint die Vegetationsdauer im Bereich der Untersuchungsfläche auffallend kurz, wenngleich die Waldbestockung zumindest während der Vegetationsperiode den extremen Einfluß der Klimafaktoren mindert.
Bodenformation des Untersuchungsgebietes
In Abhängigkeit von den Bodenbildungsfaktoren Ausgangsmaterial, Oberflächenform, Lage und Klima entwickelte sich auf der Fläche eine mehr oder weniger flachgründige Rendzina. Dieser AC-Boden, dem ein Anreicherungshorizont (B-Horizont) fehlt, besitzt wegen seiner geringmächtigen Humusauflage schlechte physikalische Eigenschaften. Diese äußern sich unter anderem in einer geringfügigen Wasserspeicherkapazität und damit großen Trockenanfälligkeit sowie in einer mäßigen Durchwurzelbarkeit. Auch seine (AC-Boden) chemischen Eigenschaften bieten keine günstigen Voraussetzungen für eine hohes Wachstum, wie die trockenrasenähnlichen Pflanzengesellschaften sowie der Buchenkrüppelwald eindrucksvoll belegen. Aufgrund der Tatsache, daß keine Bodenverlagerungsprozesse stattfinden und somit Mechanismen der Verbraunung, Stoffanreicherung und Durchmischung ausbleiben, erweist sich dieser Bodentyp als wenig wachstumsfördernde Grundlage. Zudem unterliegt der Ah-Horizont aufgrund des Standorts der Rendzina einem leichten Abtrag, so daß die Ausbildung eines mächtigen Oberbodens immer wieder unterbunden wird. Ferner trocknet, wie bereits oben erwähnt, der A-Horizont aufgrund seiner Geringmächtigkeit und der leichten Löslichkeit des Kalkes auffallend schnell aus, was eine Minderung der Aktivität der Bodenorganismen hervorruft. Dies führt zu einer Verarmung des Nährstoffgehalts, da der Prozeß der Nährstoffanreicherung in seiner Intensität erheblich herabgesetzt erscheint. Insofern weist die Rendzina nur eine geringe Variabilität in ihrer Ah-Mächtigkeit auf, was eine Unterteilung in Subtypen fraglich erscheinen läßt. Die homogene Struktur der Rendzina läßt somit nur bedingt eine Standortdifferenzierung erkennen.
Arbeitsweisen, Arbeitsverfahren und Zielsetzung der Untersuchung
Ziel der Untersuchung war es, die enorme Formenvielfalt beim Stattlichen Knabenkraut, da sie nur in wenigen der gängigen Exkursionsfloren "erwähnt" wird, zu erfassen und darzustellen. Hierzu wurden rund 500 Exemplare von Orchis mascula nach Sproßlänge, Blütenstand, Blütenfarbe, Blattanzahl, Blattspreite und Blattfarbe exakt katalogisiert, um auf diesem Wege die verschiedenen Formen bestimmen zu können. Darüberhinaus wurden die Wechselbeziehungen zwischen abiotischen und biotischen Faktoren, der belebten und unbelebten Umwelt, erörtert, indem die jeweilige Biotopstruktur in Beziehung zur Gestalt der einzelnen Orchideen gesetzt wurde. Hierzu bediente man sich einfacher Meßmethoden zur Bestimmung der Lichtqualität und Bodenstruktur. Beispielsweise wurde das BRAUN-BLANQUET-MODELL zum Bestimmen der
Pflanzenbedeckungsgrade benutzt, um den einzelnen Zonen eine Lichtqualität zuzuordnen. Mit Hilfe der Zeigerwerte nach ELLENBERG wurde das ökologische Verhalten der vorkommenden Pflanzenarten auf der Untersuchungsfläche ermittelt und daraus eine Standortanalyse abgeleitet. Diese Ergebnisse wurden im Anschluß mit den bereits gewonnenen Resultaten über die äußeren Faktoren verglichen. Daneben wurden Lichtintensität und der pH-Wert des Bodens untersucht und in Beziehung zum ökologischen Verhalten nach ELLENBERG gesetzt. Auf dieser Basis wurde es möglich, Wanderungsbewegungen und Merkmalsabstufungen herauszustellen, die wiederum Tendenzen über die weitere Entwicklung der Orchideenart zulassen. Obwohl eine genetische Untersuchung nicht durchzuführen war, konnten auf dieser Grundlage sachgerechte Vermutungen über die Entwicklung der Herkunft vorgenommen werden, und damit eine Fragestellung bearbeitet werden, die selbst heute in Fachkreisen immer noch Anlaß heftiger Diskussionen ist. Insofern sind die Aussagen am Ende der Auswertung als Hypothesen zu verstehen, die in dem einen oder anderen Fall diskussionswürdig erscheinen können. Dagegen stellen die Untersuchungsergebnisse eine solide Ausgangssituation zur Erstellung eines Maßnahmenkatalogs dar, dessen Vorschläge nicht nur die Erhaltung des Bestandes, sondern vor allem die Sicherung des Formenreichtums einer Pflanzenart auf engstem Raum zum Ziel haben.
Beschreibung der Formenvielfalt beim Stattlichen Knabenkraut (Orchis mascula)
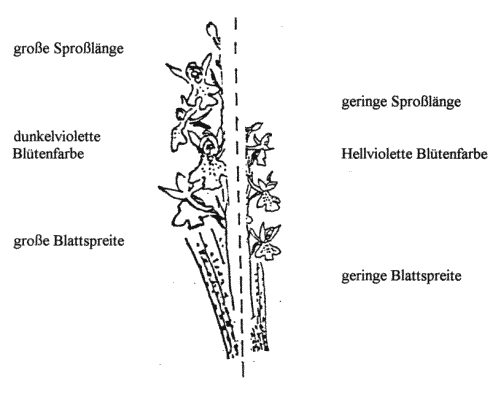
Das Stattliche Knabenkraut (Orchis mascula) zeigt auf der Untersuchungsfläche von rund zwei Hektar eine enorme Variabilität in der Gestalt (vgl. Abb. 1/Abb. 4). Die Merkmalsdifferenzierung reicht von kurzsprossigen zu langsprossigen Exemplaren. Dabei treten Unterschiede von 43 cm auf. Vor allem die Unterschiede in der Farbintensität der Einzelblüten ist hierbei auffallig. Das Farbenspektrum reicht von blaßviolett, in Einzelfallen von weißlichen Blüten, über violette Formen zu tiefvioletten dunklen Varianten. Bei einer genaueren Betrachtung der Einzelblüten findet man noch Nuancierung in der Farbgebung der Lippen. So zeigen dunkelviolette Typen oft gefleckte Lippen, während eine umforme Färbung der Kronblätter bei blaßvioletten Formen vorliegt. Erscheint der Schlund weiß, nimmt der Grad der Verblassung der violetten Farbe deutlich zu. Selbst die Gestalt der Einzelblüten läßt Variationen in der Blütenmorphologie nachweisen. Während voluminöse Einzelblüten eine markante Ausbildung des Sporns darlegen, verliert der Sporn bei kleinblütigen Formen im Verhältnis zur Gesamtblüte erheblich an Bedeutung. Die Unterlippe bei großen Exemplaren zeigt im Vergleich zu kleineren Vertretern weiter gespreizte Teillappen auf. Außerdem treten neben duftenden Formen auch duftlose Varietäten auf. In ähnlicher Weise lassen sich Formunterschiede in der Ausbildung der Blattspreite nachweise. Hierbei stehen sich lanzettliche, schmalblättrige Blätter breitflächigen Formen gegenüber. Ferner differieren die Blattfarbe und Mächtigkeit der Cuticula (Wachsschicht) erheblich. Die Blätter sind darüber hinaus in ihrer Farbgebung nochmals unterschiedlich. So sind sie entweder gefleckt oder ungefleckt. Die Flecken auf den Blättern sind bei dunkler Farbgebung meist scharf vom Farbuntergrund der Blätter abgrenzt. Bei hellgrünen Blättern erscheint die Fleckenstruktur nur schwach ausgebildet und wirkt daher eher "verfließend". Diese Blätter sind zum Teil am Boden liegend oder aufrecht stehend, die Sproßachse umfassend. Die Hochblätter variieren bei Orchis mascula in ihrer Ausformung sehr stark. Bei großen Formen erscheinen die Hochblätter rund-eiförmig, breitflächig und stengelumfassend, bei kleinwüchsigen Exemplare dagegen zurückgebildet.
Bei einer Gesamtbetrachtung der Orchis mascula-Formen lassen sich immer wieder Merkmalspaarungen oder Kombinationen in der äußeren Erscheinungsform nachvollziehen, so daß man sechs Formengruppen auf der Untersuchungsfläche bestätigen kann. Am häufigsten und in ihrer Struktur meist gegensätzlich und daher sich differenzierend gegenüber anderen Formen lassen sich dominierend auftretende Stammformen unterscheiden:
Stammformen:
- schmalblättrig
- hellgrüne Blattspreite
- kleiner Blütenstand mit vielen kleinen Einzelblüten
- duftend
- dunkelviolette Farbe der Blüte
- breitblättrig
- dunkelgrüne Blattspreite
- großer Blütenstand mi wenigen großen Einzelblüten
- nicht duftend
- blaßviolette Blütenfarbe
Auffallend häufig ist ein sogenannter "intermediärer" Typus zwischen den beiden Stammformen. Er wird durch folgende Merkmalspaarungen gekennzeichnet:
- meist breitblättrig
- kleiner, aber reichhaltiger Blütenstand, weniger gedrängt stehende Einzelblüten
- meist nicht duftend
- dunkelviolett bis violett
Neben diesen sehr häufigen und damit den Formenkreis von Orchis mascula bestimmenden Varietäten entwickeln sich Sonderformen, die die Formenfülle in beträchtlicher Weise steigern. Sie werden in ihrer Morphologie vornehmlich von den Stammformen geprägt. Hinzu kommen neue Merkmalskombinationen, die zu einer scheinbar völlig neuen Gestalt führen. Hierzu gehören vier häufig auftretende Variationen:
- schmalblättrig
- kleiner Blütenstand mit wenigen kleinen Blüten
- nicht duftend
- fast weiß bis violette Blütenfarbe
- breitblättrig
- kleiner Blütenstand mit wenigen Blüten
- nicht duftend
- dunkelviolette Blütenfarbe
- schmalblättrig
- großer Blütenstand mit wenigen großen Blüten
- nicht duftend
- hellviolette Blütenfarbe
- schmalblättrig
- großer Blütenstand mit wenigen Blüten
- duftend
- violette Blütenfarbe
Die Formenvielfalt des Stattlichen Knabenkrauts (Orchis mascula) zeigt trotz der unterschiedlichen Merkmalsausprägung eine gewisse innere Strukturierung in der Merkmalskombination auf. Obwohl bei der Trupp- bzw. Herdenbildung in nicht wenigen Fällen verschiedene Ausformungen nebeneinander existieren, bringt die räumliche Verteilung eine spezifische Konzentration einzelner Varietäten hervor.
Die Untersuchungsfläche erfahrt durch die Bestockung eine stringente pflanzensoziologische Gliederung, so daß sich drei in sich einheitliche strukturierte Zonen deutlich voneinander abgrenzen:
Zone 1 ist ein Eschenwald mit geschlossener Kraut- und Bodenschicht. Die beiden letztgenannten Stockwerke des Eschenwaldes werden in extremer Form vom einblütigen Perlgras und einigen Sauergräsern beherrscht. Die Strauchschicht beschränkt sich dagegen auf einige wenige locker gestellte Sträucher der Kahlschlaggesellschaft. Das pflanzensoziologische Gefüge weist auf eine Waldmeister-Buchen-Eschen-Gesellschaft hin. Das relativ offene Kronendach bewirkt einen starken Lichteinfall in diese Zone, was zur Folge hat, daß sich der Boden erwärmt und somit eine Austrocknung in regenschwachen Monaten leichter gegeben ist. Weiterhin fördert das offene Kronendach einen stärkeren Regeneinfall, da der Niederschlag von keinem dichten Kronendach aufgehalten und somit nicht gleichmäßiger dosiert werden kann. Die Konsequenz hieraus ist eine Lösung bzw. Auswaschung des Kalkes aus dem Boden, was zur Steigerung des von der Pflanze erreichbaren Eisengehaltes führt. Dies wird an den pH-Wert-Messungen belegt. Die pH-Werte mit 6,5 im Ah- und 6,7 im A/C-Horizont weisen auf ein schwachsaures Bodenmilieu hin (vgl. Abb. 9).
Zone 2 wird vornehmlich von einer in ihrem Zentrum dichten Edellaubholzbestockung bestimmt. Der Bestandsrand, insbesondere gegen die Eschenwaldformation, erscheint aufgerissen, was nach seiner Linienführung auf eine forstwirtschaftliche Maßnahme schließen läßt. Hier bedeckt ebenfalls eine Kraut- und Bodenschicht den Boden. Allerdings kann in diesem Fall nicht von einer Geschlossenheit durch die unteren Stockwerke gesprochen werden. Die Strauchschicht ist in dieser Waldformation nicht mehr nachzuweisen. Nach ihrer pflanzensoziologischen Struktur gehört diese Zone der Waldmeister-Buchen-Gesellschaft an.
Zone 3 wird vornehmlich von einem schwach-wüchsigen Buchenwald, dessen Kronendach geschlossen erscheint, bestimmt. Als Unterbau kommt hier die Hainbuche zur Geltung, die einen nicht unbeträchtlichen Anteil am Gesamtanteil der Holzartigen einnimmt. Vereinzelt sind den Buchen noch Eschen und andere Edellaubhölzer beigemischt, so daß die Bestockung einem Laub-Mischwald entspricht. Die Kraut- und Bodenschicht büßt erheblich an Fläche ein, wenngleich das Waldbingelkraut große Areale einnimmt. Diese Waldformation weist demnach eine Bingelkraut-Buchen-Gesellschaft aus. Das geschlossene Kronendach führt zu einer Abdunklung und Abkühlung der Zone, da das Sonnenlicht nur in Form von verstreuten Sonnenflecken in die Zone eindringen kann. Diese punktuell auftretenden Flecken erwärmen an den jeweiligen Stellen den Boden, so daß das Waldbingelkraut an diesen Stellen in geringem Maße vorkommt. Der Regen fallt in dieser Zone nicht direkt auf den Boden, da er durch das dichte Kronendach aufgehalten wird. Dadurch, daß nur kleine Wassermengen den Boden erreichen, ist nur eine geringe Kalkauswaschung in dieser Zone zu verzeichnen. Die hohen pH-Werte mit 6,95 im Ah-, 7,4 im A/C und 8,2 im C-Horizont kennzeichnen deutlich die basische Bodenreaktion.
Die spezifische Abfolge von Zone l über Zone 2 nach Zone 3, vom Eschenbestand über Edellaubholzformation zum geringwüchsigen Buchenwald bringt eine entsprechende Abstufung in der Merkmalsausbildung von Orchis mascula sowie ihrem pflanzensoziologischen Verhalten mit sich.
Wie aus Abbildung 5 zu ersehen ist, nimmt der Prozeß der Vereinzelung von Orchis mascula von Zone l nach Zone 3 drastisch zu. Formieren sich die Orchis mascula-Exemplare unter dem Kronendach der Eschen zu kompakten Trupps, die in einzelnen Arealen sogar in einer Herdenbildung mündet, so nimmt die Populationsdichte in Richtung Zone 3 erheblich ab. Obwohl im Buchenwald immer noch kleine Formationen auftreten, ist eine geschlossene Arealbildung nicht mehr nachzuweisen. Ferner findet man, im Waldbingelkraut eingestreut, in vermehrtem Maße Einzelexemplare, ein Phänomen, das sich in beiden östlich davon gelegenen Waldgesellschaften nicht belegen läßt. Zudem kann man in den Waldbingelkrautvorkommen immer wieder ungeschlechtliche Formen der Orchis mascula feststellen. Der Prozeß der Vereinzelung setzt bereits in den inneren Bereichen der Zone l ein und gelangt in der dritten voll zur Auswirkung. Es tritt infolgedessen pflanzensoziologisch auf Orchis mascula bezogen eine Verarmung in der Populationsdichte ein, so daß das Erreichen der unteren
Dichtegrenze u. a. als Ursache für das Erlöschen des Orchis mascula -Vorkommens im zentralen Bereich des Buchenbestandes anzusehen ist.
In gleicher Weise vollzieht sich die Merkmalsabstufung in der äußeren Form des Stattlichen Knabenkrauts (Orchis mascula). Vom Eschen- zum Buchenbestand nehmen Sproßlänge, Breitblättrigkeit, Chlorophyllgehalt und Spornbildung in ausgeprägter Form zu. Dagegen verhalten sich Farbintensität der Blütenblätter, Größe der Einzelblüten und Duftstoffe in umgekehrter Richtung. Diese Merkmalsdifferenzierung tritt jedoch nicht in der hier dargelegten Eindeutigkeit auf. Vielmehr kann ein Nebeneinander verschiedener Formen in einer der Zonen festgestellt werden. Insbesondere in der Zone 2 treten typische Vergesellschaftungen verschiedenster Varietäten des Stattlichen Knabenkrauts (Orchis mascula) auf engstem Raum auf, was den Schluß zuläßt, daß dieser Bereich eine Übergangszone in der Morphologie dieser Orchidee verkörpert. Doch nach den Häufigkeitsberechnungen sowie den gravierenden Abweichungen in der Merkmalsausbildung der Orchideen-Formen in der Zone l und Zone 3 ist die Feststellung einer Merkmalsabstufung gerechtfertigt. Weiterhin ist ein Vorkommen von typischen Merkmalspaarungen zu erkennen. So treten in Zone 3 z. B. viele Exemplare mit einer großen Sproßlänge, einer großen Blattspreite und einer hellvioletten Blüte auf, während in der Zone l viele Exemplare eine niedrige Sproßlänge, eine kleine Blattspreite und eine dunkelviolette Blüte besitzen. Auch hier ist wieder eine Abstufung zu erkennen (vgl. Abb.6).
Interpretation der Auswertung
Die Umweltfaktoren und mit ihnen die jeweiligen Pflanzengesellschaften prägen in entscheidendem Maße die abiotische Struktur. Vor allem Licht-, Temperatur- und Bodenverhältnisse stellen dabei die entscheidenden Faktoren dar, die als abiotische Selektionsfaktoren maßgeblich für die Formenvielfalt verantwortlich sind.
Standortansprüche des Stattlichen Knabenkrauts (Orchis mascula) lassen sich nach der pflanzensoziologischen Exkursionsflora nach OBERDORFER und der Exkursionsflora nach ROTHMALER wie folgt definieren:
Orchis mascula kommt häufig in Halbtrockenrasen (Mesobrometum), auch in Eichen-Hainbuchen-Wäldern sowie in nährstoffreichen Eschen-Buchenwaldungen vor. Die zuletzt genannte Waldformation mit ihren Abwandlungen und Übergängen entspricht der Untersuchungsfläche. Die als Halbschatten-Lichtpflanze charakterisierte Orchideenart bevorzugt mäßig trockene bis frische, mehr oder weniger nährstoff- und basenreiche, dafür aber oft kalkarme, aber mild bis mäßig saure, humose, mittel- bis tiefgründige Lehmböden. Schon bei einer Bewertung der hier dargelegten Standortansprüche wird offenkundig, daß Orchis mascula bei den abiotischen Faktoren eine gewisse Toleranz besitzt.
Stellt man diesen Aussagen das in Form von Zeigerwerten nach ELLENBERG festgelegte ökologische Verhalten der Orchis mascula gegenüber, so erhält man für Licht-, Temperatur-, Kontinental-, Feucht-, Reaktions- und Stickstoffzahl folgende Werte:
|
Lichtzahl Temperaturzahl Kontinentalzahl Feuchtzahl Reaktionszahl Stickstoffzahl |
7 X 3 4 8 3 |
Diese Werte für die spezifischen Umweltfaktoren weisen das Stattliche Knabenkraut (Orchis mascula) als Halblichtpflanze aus, die sowohl im vollen Licht als auch im Schatten gedeihen kann. Während sie hinsichtlich der Temperatur eine weite Spanne aufweist, also ein indifferentes Verhalten zeigt, bevorzugt sie ein ozeanisch bis subozeanisches Klima, was ihr eine Verbreitung in großen Teilen Mitteleuropas ermöglicht. Ihre Feuchtzahl kennzeichnet ebenfalls eine Mittelstellung. Dabei steht sie zwischen Trocknis- und Frischezeigern, was ihr die Fähigkeit bescheinigt, sowohl auf trockenen als auch auf mittelfeuchten Böden wachsen zu können. Auch diese abiotische Raumstruktur liegt im Untersuchungsgebiet vor. Zum einen stockt der Eschenbestand (vgl. Zone 1) auf einer frischen bis mittelfeuchten Rendzina, zum anderen kann sie sich auch noch auf trockenen Böden behaupten. Gerade im Bereich der Zone 2 und 3 neigen die eher flachgründigen, wenig humosen, allerdings mit Laubstreu bedeckten Rendzinen bei entsprechender Witterung zur Austrocknung, so daß vor allem die
Vegetation der Kraut- und Bodenschicht sehr schnell unter Wassermangel leidet. In ähnlicher Weise läßt die Reaktionszahl ein gewisses Schwanken in ihrem ökologischen Verhalten hinsichtlich der Bodenreaktion und des Kalkgehaltes erkennen. Der Toleranzbereich von Orchis mascula reicht speziell in diesem Fall von Schwachsäure- bis Schwachbasenzeiger und bestätigt ferner das Vorhandensein von Kalk als Ausgangsgestein. Der bankig verwitterte, oberflächennah anstehende Kalk bestätigt für das gesamte Untersuchungsgebiet in zutreffender Weise die Aussage- und Interpretationsfähigkeit der Zeigerwerte für das Stattliche Knabenkraut (Orchis mascula). Dagegen legt seine Stickstoffzahl eine enge ökologische Amplitude fest, die ihr Vorkommen vornehmlich auf stickstoffarmen Standorten charakterisiert. Auf mittelmäßigen bis stickstoffreichen Böden wird demnach ihr Wachstum sichtbar erschwert.
Stellt man die von ELLENBERG, ROTHMALER und OBERDORFER definierten Standortansprüche denen der Untersuchungsfläche gegenüber, kommt man zu einer Übereinstimmung in den wesentlichen Kriterien des Standorts für Orchis mascula. Hierbei tritt bei nahezu allen Angaben eine hohe Variabilität in den jeweiligen Standortansprüchen auf. Sie läßt sich insbesondere durch die Dreigliederung der Untersuchungsfläche transparent nachweisen. Aufgrund dieser Tatsache basiert auch der hohe Modifikationsgrad (nicht erblich, sondern durch äußere Einflüsse bedingte Veränderung) bei Orchis mascula, der sich in extremer Weise in dem Formenreichtum innerhalb der Orchideenart niederschlägt.
Aufgrund des geschlossenen Kronendaches im Buchenbestand ist die Intensität des Tageslichtes herabgesetzt, und die daraus folgende Tendenz zum Längenwachstum bedingt eine morphologische Veränderung der Pflanzengestalt. Es tritt die charakteristische Erscheinung der Vergeilung, auch Etiolement genannt, auf. Auch wenn bei den Einkeimblättrigen wie beim Stattlichen Knabenkraut (Orchis mascula) weniger die Länge der Sproßachse als vielmehr die Länge der Blätter verändert wird, so weist das Längenwachstum von Orchis mascula deutlich auf einen Lichtmangel hin. Es findet also durch die Veränderung der abiotischen Faktoren, bezogen auf den Standort der Orchideen, eine Umwandlung von Blättern und Sproß statt. Anhand der Abbildung 3 wird offenkundig, daß die Veränderung der äußeren Gestalt eine Reaktion auf einen zu geringen Lichteinfluß darstellt. Die typischen Etiolierungserscheinungen, wie kleine bis rückgebildete Blattspreiten mit bleichem gelblichen Aussehen, was auf ein Unterbleiben der Chlorophyllbildung zurückzuführen ist, kann in dieser Form nicht bestätigt werden. Die Zunahme der Blattspreite in und der damit hergehende Anstieg des Chlorophyllgehaltes weist vielmehr auf den mesomorphen Charakter von Schattenblättern hin. Hingegen zeigen die Blätter in Richtung Zone l eher die Morphologie eines mesomorphen-skleromorphen Typus auf, was auf eine lang anhaltende intensive Sonneneinstrahlung zurückzuführen ist. Die Aussage wird darüber hinaus durch die Tatsache unterstützt, daß glänzende Formen in Zone l und vor allem in Zone 2 vorkommen. Es handelt sich dabei um eine Steigerung der Wachsschicht mit der funktionalen "Zielsetzung", die Verdunstung herabzusetzen.
Wenn auch die Eindeutigkeit im Sinne einer Merkmalsabstufung nicht immer erkennbar ist, weil in Zone 3 in Teilarealen die glänzenden Formen überwiegen, so kann dieses als Ergebnis der Auswirkung von Sonnenflecken verstanden werden. Nach der BLACKMAN-Regel ist zu sagen, daß die Variabilität in enger Beziehung zu einzelnen äußeren Faktoren oder zu dem im Maximum befindlichen äußeren Faktor steht. Die Intensität der Blütenfarbe nimmt von Zone l zu Zone 3 ab (vgl. Abb. 1), das bedeutet, daß es in Zone l überwiegend dunkelviolette Exemplare und in Zone 3 überwiegend violette bis hellviolette Exemplare gibt. Dies ist durch den unterschiedlich großen Kalk- und Eisengehalt des Bodens zu erklären, da ein hoher Eisengehalt die Farbintensität erhöht. In Zone l ist der Kalkgehalt geringer, so daß die Pflanzen eher an Eisenionen gelangen können. In der Zone 3 ist durch die geringere Kalkauswaschung ein höherer Kalkgehalt zu verzeichnen. Somit wird die Möglichkeit der Eisenaufnahme der Pflanze verringert. Die Farbintensität nimmt folglich ab.
Es liegt eine ökologische Merkmalsverschiebung vor, entlang allmählich sich verändernden Umweltfaktoren (vgl. Abb.3/Abb. 6). Die Merkmalsdifferenzierung erfolgt kontinuierlich. Selbst unter der Berücksichtigung, daß genetische Untersuchungen nicht angestellt wurden und das Untersuchungsgebiet als kleinräumig anzusehen ist, kann man von einer Cline-Bildung (stufenlose Merkmalsverschiebung) sprechen. Trotz der relativ scharf abgesetzten
Standorte und Lebensräume und der Merkmalsdifferenzierung zwischen Zone l und 3 kann man nicht unbedingt von einer Unterscheidung nach ökologischen Rassen sprechen. Die Merkmalsabstufung ist nämlich fließend und ist nicht deutlich genug abgestuft. Die spezifische Merkmalsbildung, die bei den Orchideen des Eschenbestandes von denen des Buchenbestandes divergiert, lassen auf Wirkungsweisen äußerer Selektionsfaktoren schließen. Insgesamt kann man dennoch von einer ökologischen Differenzierung des Artbereiches durch eine Öko-Cline-Bildung (... rein auf morphologischer Basis) sprechen, da verschiedene Funktions- und Merkmalsbereiche erfaßt wurden. Sie ist also modifikatorisch bedingt, d. h., daß Pflanzen mit völlig gleichem Erbgut unter scheinbar gleichen Bedingungen eine Variabilität aufweisen. Aufgrund der Merkmalsverschiebung weist Orchis mascula eine fließende Modifikabilität auf, bei der die Modifikationen fortlaufend gradweise Veränderungen eines Merkmals aufweisen. Dies wird vor allem mit der Zunahme der Sproßlänge und der Nuancierung der Farbintensität offenkundig. Nach dem Vorkommen der hier bestimmten Formenvielfalt ist nicht auszuschließen, daß unter gewissen Umständen die Modifikation erhalten bleibt. Aufgrund der clinalen Differenzierung ist eine Durchmischung des Genpools (Gesamtheit aller Gene innerhalb einer Population) gegeben. Damit ist eine Aufspaltung der Art Orchis mascula in ökologische Rassen nicht zutreffend. Jedoch beim Nachweis von Modifikationen -nicht unter der Voraussetzung lang anhaltender modifikatorischer Wirkung besteht unter Umständen die Möglichkeit, insbesondere durch die Merkmalsunterschiede der Orchideen in Zone l und Zone 3, daß diese Modifikationen sich auf die Art auswirken, indem Mutationen (genetische Veränderung) zur Wirkung gelangen und somit zur Aufspaltung der Art in Rassen führen. Der hier beschriebene Weg der Rassenbildung kann aber nur dann gewährleistet sein, wenn die räumlich stark abgegrenzte Annidation möglich ist. Da davon auszugehen ist, daß die Familie der Orchidaceen -in ihrer Entwicklung- sehr jung ist und eine typische schrittweise Merkmalsverschiebung schneller zur Wirkung kommt, herrscht im Fall des Untersuchungsgebietes eine sich verändernde Auslese vor, die nur dann in eine stabilisierende übergehen kann, wenn die drei unterschiedlichen ökologischen Nischen über einen langen Zeitraum erhalten werden und die nachgewiesene modifikativ bedingte Differenzierung sich auf die Genstruktur auswirkt und somit den Weg zur Rassenbildung darstellt.
Aufgrund der hohen Populationsdichte des Stattlichen Knabenkrauts (Orchis mascula), was die Herdenbildung im Bereich des Eschenbestandes belegt, ist die Annahme gerechtfertigt, daß diese als Ausgangs- bzw. als Ausgangspopulation für die Ausbreitung der Orchideenart gewertet werden kann (vgl. Abb. 5). Von dieser Population aus ist es einzelnen Exemplaren gelungen, in die Zonen 2 und 3 zu wandern, wobei eine typische Kanalisierung parallel zum Hang erfolgt zu sein scheint. Doch darf die strenge Wanderungsrichtung nicht absolut gesehen werden. Vielmehr gelingt es Orchis mascula, vom Zentrum des Pools ausgehend sich in alle Richtungen kreisrund auszubreiten. Allerdings wirken in der Peripherie einzelne abiotische Faktoren und unter diesen vor allem die Lichtqualität und -quantität in einer Art und Weise, daß die kreisähnliche Streuung einzelner Teilpopulationen eingedämmt wird, was der Prozeß der Vereinzelung, der schließlich in die Vereinzelung endet, untermauert. Diese Gegebenheit trifft für alle drei Zonen des Untersuchungsgebietes zu.
Bei einer oberflächlichen Betrachtung scheint der Vorgang der Streuung zur Wirkung zu kommen, der jedoch nicht unweit vom Zentrum der Population zum Erliegen kommt. Insofern erscheinen die Wanderungsrichtungen in nördlicher und südlicher Richtung mäßig zu sein, während die West-Ost-Migration von höchster Intensität ist, was sowohl die Hauptwanderungsrichtung als auch die Bildung von Teilpopulationen in Zone 2 und 3 belegen. Die Lebensfähigkeit der Teilpopulation ist jedoch in bezug auf eine Herdenbildung in der Edellaub- und Buchenwaldzone gering, wenngleich Einzelexemplare höchste Wachstumsintensitäten erkennen lassen. Diese Annahme ist insofern haltbar, wenn man bedenkt, daß von den großen Orchis mascula - Formen keine deutliche Ausbreitung erfolgt. Sie besitzen demnach nicht die Fähigkeit, als Ausgangsbasis für eine Ausdehnung der Art dienen zu können. Lediglich im Bereich der Eschenwaldzone spalten sich kleinere Areale der Orchideenart vom Zentrum ab. Dies beruht sowohl auf der einheitlichen Struktur der Fläche, als auch auf der Populationsdichte im Zentrum der
Initialpopulation. Andererseits ist es auch denkbar, daß es im Laufe der Entwicklung der Waldformation zu einer Auflösung der Gesamtpopulation gekommen ist, so daß es hier das Phänomen einer Streuung gegeben haben könnte. Durch das Einsetzen des Kronenschlusses bei einem selbst schwachwüchsigen Buchenkrüppelwald tritt eine Veränderung in der äußeren Struktur des gesamten Areals ein. Eine Verringerung der Lichtverhältnisse und eine Vereinheitlichung der klimatischen Faktoren verändern die standörtlichen Gegebenheiten im Mikrobereich. Insofern ist die Annahme einer Auflösung des Gesamtbestandes denkbar. Nach der Merkmalsdifferenzierung, die das Stattliche Knabenkraut (Orchis mascula) als formenreiche Art ausweist, handelt es sich auf der Untersuchungsfläche um Teilgesellschaften, die als Standortmodifikationen zu bewerten sind (vgl. Abb. 3). Es handelt sich somit um eine Anpassung an die standörtlichen Gegebenheiten.
Dennoch belegen die Merkmalsdifferenzierungen eine einsetzende Formenvielfalt, die je nach naturräumlicher Ausstattung zur Entwicklung stabiler Teilpopulationen führen kann. Die Variabilität liegt demnach unterhalb der Unterart, also nicht im Bereich der Sub-species, sondern im Bereich der Varietätenausbildung. Es ist also immer noch ein einheitlicher Genpool vorhanden.
Bei langanhaltenden ähnlichen äußeren Einflüssen kann es möglicherweise zu einer Stabilität der Formen kommen, so daß eine starke Unterscheidung der Teilpopulationen in Erwägung zu ziehen ist.
Taxonomie und Systematik
Die Stattliche Knabenkraut (Orchis mascula) -Population zeigt auf engstem Raum innerhalb des Untersuchungsgebietes eine Formenvielfalt, die zunächst auf eine Aufspaltung des Genpools schließen läßt. Doch die stufenweise Merkmalsdifferenzierung in Form einer Cline-Ausbildung belegt eindeutig, daß der Genfluß zwischen den einzelnen Teilpopulationen aufrechterhalten bleibt. Somit ist die freie Durchmischung des Genpools nicht eingeschränkt, eine freie Kombinierbarkeit (Rekombination) ist daher gewährleistet. Die Teilpopulationen von Orchis mascula sind aufgrund ihrer Variabilität als offene Systeme
zu bezeichnen. Dennoch weisen die Formen der Zone l und 3 bereits deutliche Unterschiede in ihrer Morphologie auf, was einerseits die These einer Öko-Cline-Bildung stützt, andererseits stehen hierzu die Anzahl der Zwischenformen in Zone l und 2 sowie die geringe geographische Distanz mit ihrer Funktion einer geographischen Separation im Widerspruch zur obigen Annahme. Bei einer Bewertung aller Kriterien hinsichtlich der Darstellung der evolutiven Entwicklung der Teilpopulationen (=Deme) von Orchis mascula ist davon auszugehen, daß der Genfluß durch Wanderung nur wenig behindert ist. Da selbst die Distanzisolation gering erscheint und somit kaum ein Gefälle der Allelhäufigkeiten angenommen werden kann, ist die Annahme einer Cline-Bildung fraglich. Insofern ist es anzuzweifeln, ob die Trennung der Orchis mascula -Formen stark genug ist, um Lokalrassen (Subspecies) auszubilden.
Mit Sicherheit reichen die Merkmalsunterschiede jedoch aus, um die Deme als Varianten einer Art definieren zu können. Ferner steht fest, daß bei der Wanderung verschiedener Orchis mascula -Individuen in verschieden strukturierten ökologischen Nischen, die spezifisch durch die pflanzensoziologische Struktur bestimmt werden, Umwelteinflüsse zu Abwandlungen ihrer Morphologie geführt haben. Diese Anpassung an verschiedene Standorte ruft im Rahmen der ökologischen Einnischung eine unwahrscheinliche Formenfülle hervor, die eine eindeutige Zuweisung der Individuen zu ein und derselben Rasse sichtbar erschwert. Hierbei handelt es sich um eine fließende Modifikabilität, bei der die Modifikationen eine fortlaufende gradweise und stetige Formabwandlung hervorrufen. Da in den einzelnen Zonen verschiedene Formtypen gleichzeitig auftreten können, sogar in einem Areal einer Zone nebeneinander gedeihen, herrscht eventuell sogar die alternative Modifikabilität. Betrachtet man jedoch die Merkmalsabstufung in den drei Vorkommen, ist der erstgenannten Modifikabilität der Vorrang einzuräumen. Wenn man den allgemeinen Gesetzmäßigkeiten der Modifikation folgt, daß genetisch festliegt, welche Gene durch Umweltfaktoren beeinflußt werden können und auf welche Art und Weise die Pflanzen auf die gegebenen Umweltbedingungen reagieren, dann steht außer Frage, daß eine Vererbung erworbener Eigenschaften nicht gegeben ist. Es könnte lediglich bei konstant divergierenden Umweltbedingungen zu einer
Stabilisierung der Modifikation mit nachfolgender Mutation kommen, die schließlich zu einer Rassenbildung fuhrt. Um endgültigen Aufschluß über die Einordnung der formenreichen Art befriedigend lösen zu können, bedarf es verschiedener genetischer Untersuchungen, die im Rahmen dieser Untersuchung nicht zu leisten waren. Es handelt sich aber aller Wahrscheinlichkeit nach bei den untersuchten Demen von Orchis mascula um Standortmodifikationen. Da verschiedene Formen von Orchis mascula in einzelnen Herden gemeinsam auftreten, befindet sich die Population in der Aufspaltung unter dem Niveau der Lokalrassen.
Maßnahmen zur Erhaltung des Bestandes
Aufgrund der Einzigartigkeit des Stattlichen Knabenkraut (Orchis mascula) -Bestandes, die sich in der Populationsdichte und im Formenreichtum äußert, sind folgende Maßnahmen zur Erhaltung zu beachten:
I. Die Struktur der drei ökologischen Nischen - bestehend aus mittelaltem Eschenbestand, Edellaubholz und Buchenaltholz mit Edellaubhölzern durchsetzt und mit der Hainbuche als Unterholz - ist zu erhalten, um die unterschiedliche Standortstruktur aufrechtzuerhalten, denn nur auf diese Weise ist der hohe Grad der Standortmodifikation bei Orchis mascula zu gewährleisten
II. Forstliche Maßnahmen sollten behutsam vorgenommen werden:
A. Die Erhaltung des Eschenbestandes ist dringend geboten.
B. Eine Erweiterung des Edellaubholzbestandes durch eine gezielte Auflockerung in Richtung Buchenaltholz ist unter Umständen zu befürworten, damit durch eine Erweiterung der Übergangszone die Möglichkeit einer weiteren Verbreitung der Orchideen gegeben ist.
C. Eingriffe in das Buchenaltholz sollten insbesondere bei Kronenschluß vorgenommen werden, um die abiotischen Voraussetzungen für das Wachstum des stattlichen Knabenkrauts zu sichern. Hierbei erscheint eine selektive Entnahme einzelner Baumstämme empfehlenswert.
III. Bei Bringung des eingeschlagenen Holzes sind Rückeschäden, Bodenverdichtungen und Fahrzeugspuren zu vermeiden.
IV. Der Holzeinschlag sowie die Bringung des Holzes aus dem Bestand sollte in der vegetationsfreien Zeit erfolgen. Auf diese Weise können Beschädigungen der Pflanzen ausgeschlossen werden.
V. Bei starker Durchfeuchtung des Bodens sollte vor allem das Rücken des Holzes mit schweren Maschinen unterbleiben, um Bodenverdichtungen zu vermeiden.
VI. Das Rücken der Stämme empfiehlt sich schonend mit Hilfe von Pferden vorzunehmen.
VII. Das Areal sollte den Waldbesuchern nicht zugänglich gemacht werden, da ansonsten die Gefahr von Trittschäden gegeben ist.
VIII. Eine Eingatterung des Gebietes ist empfehlenswert, um Wühlschäden durch Schwarzwild und Dachs zu vermeiden. Der Zaun ist in diesem Fall in den Boden einzugraben. Ferner sollte zur Unterstützung der Schutzmaßnahmen der Jagddruck in der Umgebung der Untersuchungsfläche verstärkt werden.
IX. Pflegemaßnahmen im Sinne der Vermeidung eines möglichen Konkurrenzdrucks durch Perlgras oder Waldbingelkraut sind nach Abschätzung der Sachlage möglich, jedoch nach dem derzeitigen Entwicklungsstand nicht erforderlich. Sollte allerdings der erstgenannte Fall eintreten, sind die Eingriffe in die Krautschicht keineswegs flächendeckend vorzunehmen. Die Durchführung sollte dabei manuell erfolgen.
X. Bei einem verstärkten Auftreten von Sträuchern infolge einer Auflichtung der Bestände sollte bei hohem Bedeckungsgrad der Kahlschlaggesellschaften diese auf ein gewisses Maß zurückgedrängt werden, damit der Konkurrenzdruck eingeschränkt wird.
XI. Bodenverändernde Vorgänge oder Bodenverwundungen, die vor allem den Ah-Horizont betreffen, sind zu unterlassen, um die Symbiosen nicht zu beeinträchtigen
XII. Die Bestände sollen gegenüber den angrenzenden Waldungen unbeeinflußt bleiben, damit eine indirekte Einflußnahme von außen auf die Orchideenpopulation ausbleibt.
Literaturnachweis
Ellenberg, Heinz (1982): Vegetation Mitteleuropas mit den Alpen. Stuttgart.
Ellenberg, Heinz (1992): Zeigerwerte von Pflanzen in Mitteleuropa. Göttingen.
Kühn, A. (1984): Grundriß der Vererbungslehre. Heidelberg.
Oberdorfer, E. (1983): Pflanzensoziologische Exkursionsflora. Stuttgart.
Rothmaler, Werner (R. Schubert/W. Vent) (1986): Exkursionsflora. Berlin.
Runge, F. (1990): Pflanzengesellschaften Mitteleuropas. Münster.
Thommen, Eduard (1973): Taschenatlas der Schweizer Flora. Stuttgart.
Walter, Heinrich (1973): Allgemeine Geobotanik. Stuttgart.
Winkler, Sieghard (1979): Einführung in die Pflanzenökologie. Ulm.
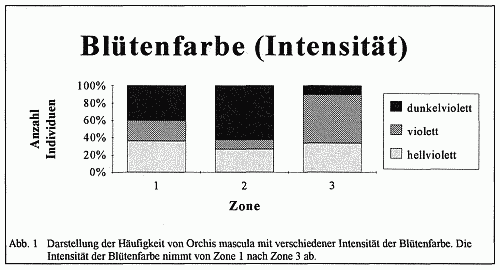
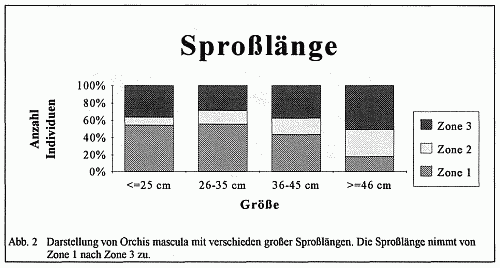
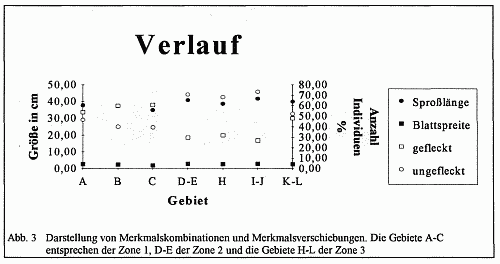
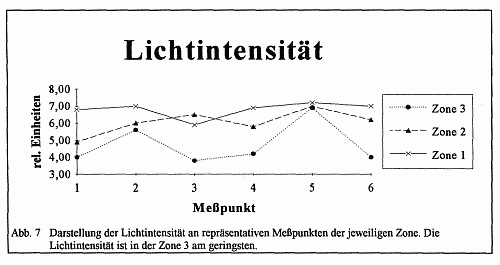
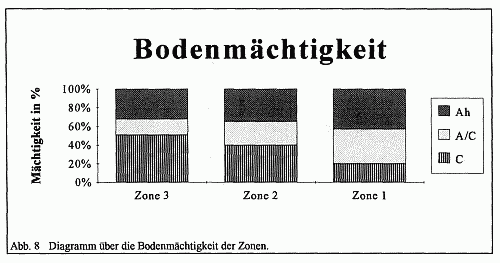
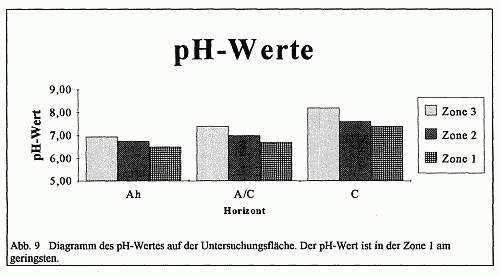
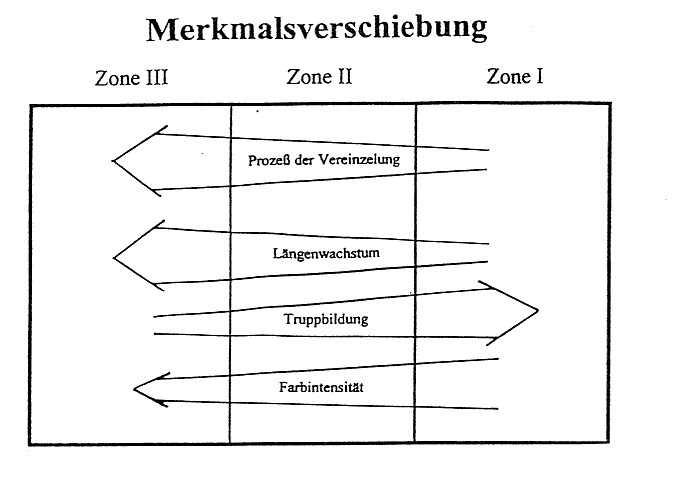
Abb. 4 Die Merkmalsverschiebung der einzelnen morphologischen Aspekte
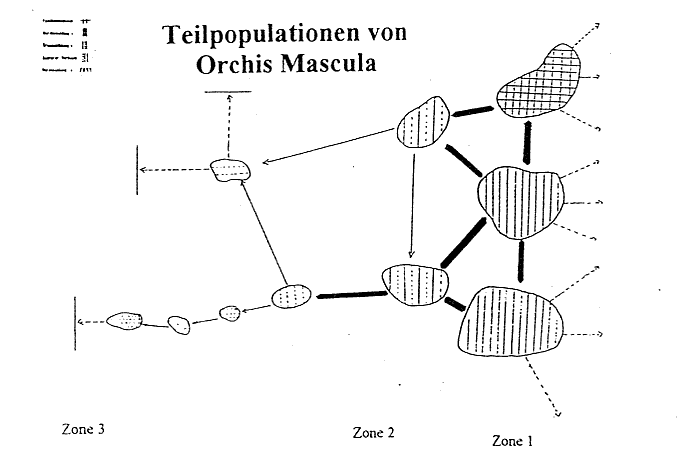
Abb. 5 Grobe Kartierung der verschiedenen Teilpopulationen von Orchis mascula mit Migrationspfeilen (Hypothese)
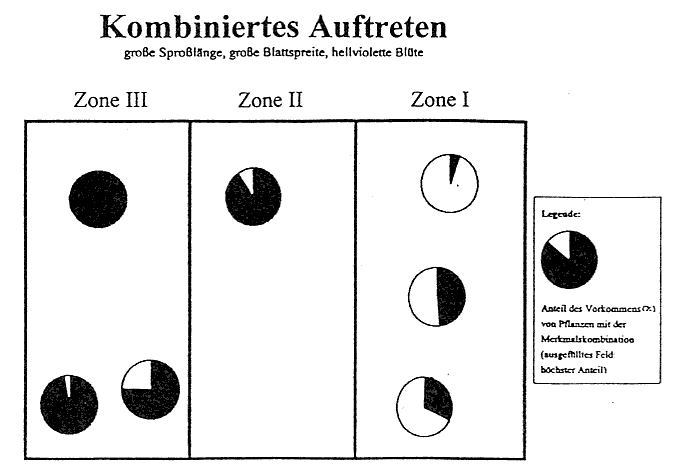
Abb. 6 Das kombinierte Auftreten von Orchis mascula mit den morphologischen Eigenschaften "große Sproßlänge, große Blattspreite, hellviolette Blütenfarbe".
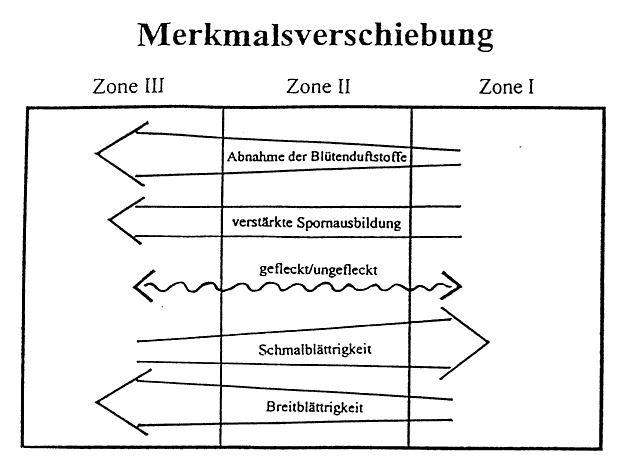
Abb. 4b Die Merkmalsverschiebung der einzelnen morphologischen Aspekte
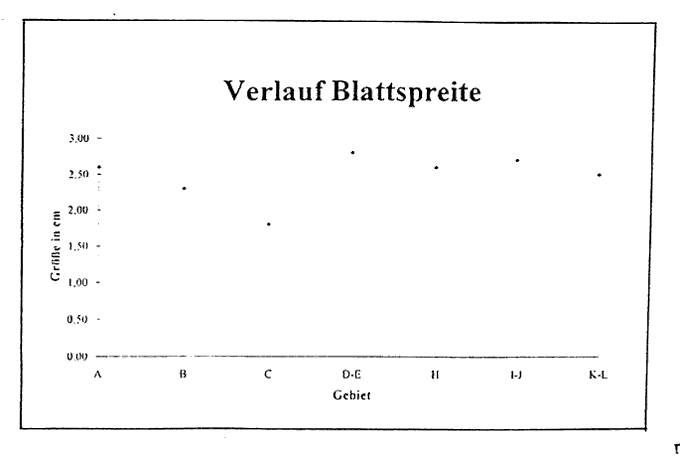
Abb. 10 Die Größe der Blattspreite in den jeweiligen Gebieten. Die Gebiete A-C entsprechen der Zone 1, D-E der Zone 2 und H-L der Zone 3.
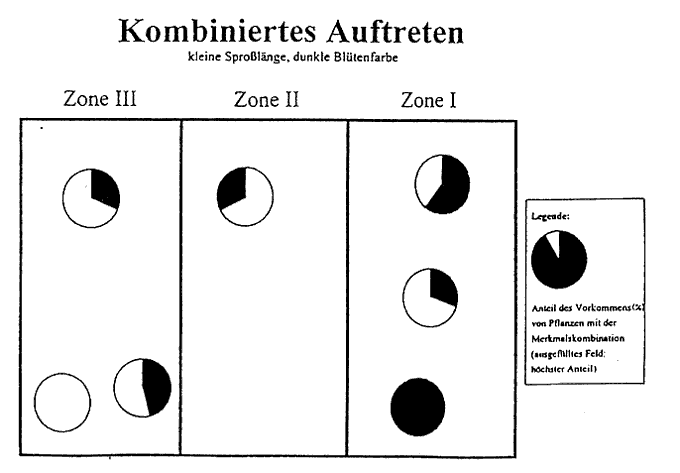
Abb. 6b Das kombinierte Auftreten von Orchis mascula mit den morphologischen Eigenschaften "kleine Sproßlänge, dunkelviolette Blütenfarbe".