| EGGE-WESER | Bd. 7 | Seite: 57-78 | Höxter 1995 |
Die "Grundlosen" bei Höxter
Ein Beitrag zur Vegetations- und Siedlungsgeschichte der Umgebung von Höxter
Fritz-Rudolf Averdieck und Kurt Preywisch
Zusammenfassung
Bei den (1956 erkennbaren) zehn Hohlformen des ND Grundlosen bei Höxter handelt es sich sehr wahrscheinlich um Dolinen über Gips- oder Salzauslaugungen in geringerer oder größerer Tiefe des Untergrundes. Nachweislich seit 1467 wurden sie als Wasserlöcher beschrieben. Erhebungen über die Anteile der Vegetationselemente ermöglichte erst der Urkataster in Preußen ab 1830. Auch die Artenzusammensetzung der Wälder und Forsten findet erst spät Erwähnung. Bis in das 18. Jahrhundert werden nur Eichen, Buchen und "anderes Holz" genannt.
Die Waldzerstörung gibt spätestens um die Mitte des 14. Jahrhunderts Anlaß zu (meist wirkungslosen) Gegenmaßnahmen. Der Ziegenberg, seit 1365 namentlich so geführt, trägt als Folge von Überweidung noch heute Reste der Wachholderheide.
In unmittelbarer Nähe der Grundlosen wird zwischen 1357 und 1495 Hopfen genannt, in ihrem Bereich selber in der 2. Hälfte des 15. Jahrhunderts Weidenbruch zum Rutenschneiden. Die Vegetation an und auf den Grundlosen hat sich in unserem Jahrhundert infolge menschlicher Eingriffe unter Artenschwund stark verändert.
Palynologische Untersuchungen wurden an einem Bohrprofil vorgenommen, das bei 815 cm das Holozän wegen Steinanreicherung nicht durchteufen konnte. Bruchwaldtorf oder Grobdetritus-Gyttja wechseln mit Ton- und Muddeschichten, besonders im unteren Teil, mehrfach. Die Folge der neben- oder übereinander aufgewachsenen Verlandungsgesellschaften geht aus den pflanzlichen Großrestfunden (Tab. 2) und den Pollenanalysen hervor.
Im Pollendiagramm sind sämtliche Kurven auf die Summe der Waldbäume unter Ausschluß der dominierenden Erle und Weide bezogen. Die Summenkurven der unbedeutenderen oder selteneren Arten, Gattungen oder Familien sind in Tab. l aufgeschlüsselt.
Das Diagramm ist in die zehn Abschnitte a bis k untergliedert und beginnt mit dem ausgehenden Atlantikum. Auf eine EMW-Haselzeit mit reichlich Linde vollzieht sich unvermittelt mit dem Übergang zum Subatlantikum der gleiche Anstieg und die ähnliche Dominanz der Buche wie in den Diagrammen der benachbarten Solling-Hochmoore. Zeitweilig miteinander vikariierende Kurvenschwankungen von Eiche, Buche und Hainbuche, zeitweilig auch von der Hasel, stehen im Zusammenhang mit dem Wechsel der Siedlungsanzeichen seit der vorchristlichen Eisenzeit. Mit deren starkem Anstieg spätestens ab dem 6. oder 7. Jahrhundert n. Chr. erleiden die beiden letztgenannten Bäume starke Einbußen zugunsten der Eiche.
Nadelholzpollen erlangen erst in den obersten Spektren Bedeutung. Während sich im unteren Diagrammteil keine größeren Siedlungsunterbrechungen abzeichnen (bei allerdings großen Probenabständen), kommt am Ende der Frühen Eisenzeit eine starke Siedlungsrezession zum Ausdruck, die bis in die Zeit zwischen dem 4. und 6. Jahrhundert anhält (Tab. 3).
Durch gezielten Ansatz von 14C-Datierungen sind im unteren Profilteil 2 rasch sedimentierte, sich invers überlagernde Abschnitte um 1100 bezw. 600 v. Chr. auszumachen, die auf Sedimentrutschungen schließen lassen und damit die Erdfall-Hypothese stützen.
Weitere Untersuchungen aus dem Bereich der Grundlosen an verschiedenem Rezentmaterial deuten auf früher stärkere forst- und landwirtschaftliche Nutzung der Umgebung.
Danksagung
Herr Dr. H.-J. BRÜNING (Höxter) half bei den archivalischen Studien. Archäologische Hinweise gab Herr Dr. H.-G. STEPHAN (Göttingen). Gerätetransport, Sondierungen und Bohrungen mußten an einem Wochenende vorgenommen werden. Sie wären ohne das Engagement der damaligen Studenten Frau Heidemarie AVERDIECK und Herr Dietrich GOTHE (beide Kiel) kaum möglich gewesen. Das Livingstone-Kernbohrgerät konstruierte, baute und betreute Herr Ing. Robert GISEWSKY (Plön). Die Pollen- und Makroanalysen führte Frau M. NEVE (Kiel) aus. Die Moose bestimmte Herr Dr. P. M. KLINGER (Oldenburg). Allen Genannten sei für ihre Hilfe herzlich gedankt
Die Vegetationsentwicklung in historischer Zeit (Preywisch)
Einleitung
Im Wesertal liegt knapp 2 km südlich von Höxter bei 51° 45' N und 9° 22' E in einer Erstreckung von wenig über einem halben Kilometer schrotschußförmig verteilt eine Gruppe von rundlichen Eintiefungen (Abb. l u. 2). Drei von ihnen waren in den meisten der letzten Jahre mit Wasser gefüllt. Seit 1974 sind sie vom flächigen Naturdenkmal "Grundlosen" eingeschlossen. Die Verordnung (Regierungsbezirk Detmold vom 17. 2. 1975, Nr. 7) enthält keine nähere Begründung. Die schon viele Jahre vorher vom Verf. betriebene Unterschutzstellung gewann erst Gestalt nach einer Tagungsexkursion des Westfälischen Naturwissenschaftlichen Vereins und einer vom ersten Vorsitzenden, Herrn Dr. Runge, verfaßten Resolution vom 26. 5. 1972 an den Bürgermeister von Höxter. In diesem Brief heißt es u. a.: Die Tagungsteilnehmer haben bei ihrer Exkursion in der Weseraue mit Erstaunen den Reichtum der Grundlosen in pflanzensoziologischer, floristischer und tiergeographischer Hinsicht festgestellt. So wurden . . . das äußerst seltene Moorkreuzkraut (Senecio tubicaulis) und der Seefrosch (Rana ridibunda) entdeckt", der bisher in ganz Westfalen nur hier beobachtet wurde (vgl. RAUS 1977).
Die Floristisch-Soziologische Arbeitsgemeinschaft führte auf ihrer Tagung 1976 ebenfalls eine Exkursion in dieses Gebiet. Angesichts der Bedrohung seiner Ökologie durch den nahen Kiesabbau wurden auf der Abschlußdiskussion eingehendere Untersuchungen zur Flora und Fauna, auch hinsichtlich ihrer Entstehung, beschlossen.
Im vorliegenden Beitrag legen die beiden Verfasser die Ergebnisse ihrer Nachforschungen zur Entwicklung der Grundlosen und der Vegetation der engeren Umgebung in geschichtlicher und vorgeschichtlicher Zeit vor. Zur Mitarbeit konnte ein weiterer Autor gewonnen werden, der durch die absoluten Datierungen ein Zeitgerüst beisteuerte, das die Erkennung von Komplikationen möglich machte (s. Seite 79 ff).
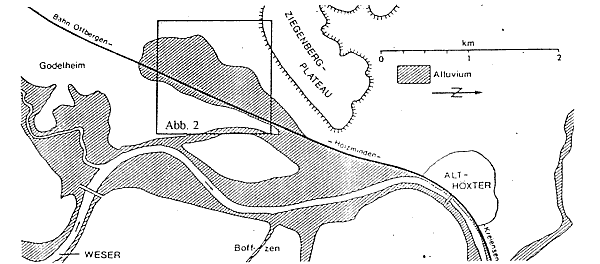
Abb. 1: Das „Alluvium“ bei Höxter, n. GRUPE (1927).
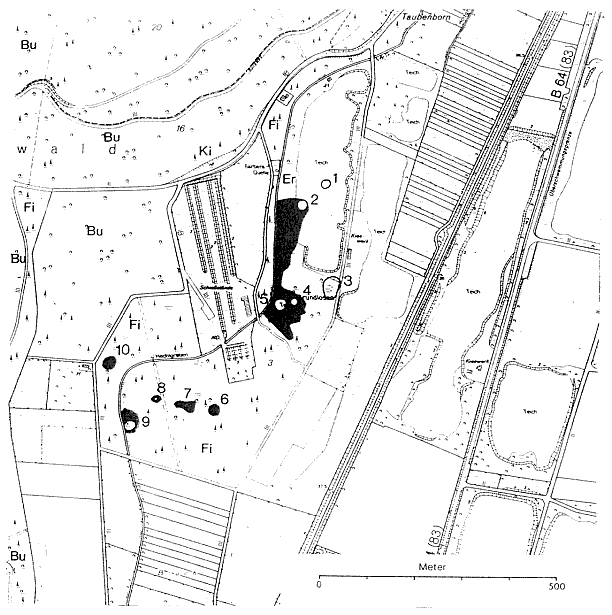
Abb. 2: Die Grundlosen und ihre nächste Umgebung aus der Deutschen Grundkarte von 1982 (vervielfält. mit Genehmigung des Kreises Höxter - Kataster- u. Vermessungsamt - v. 07.11.85 Nr. 282/85). Darin sind zusätzlich eingetragen die Wasserflächen (Umriß) und die Feuchtvegetation (Raster) von 10 Grundlosen, welche in einer Luftbildkarte von 1956 noch erkennbar waren. Außerdem sind die Hauptbaumarten nach der Forstkarte der Stadt Höxter vom 01.10.76 vermerkt.
Topographie und Toponomastik
Auf einer Luftbildkarte von 1956 sind zehn Eintiefungen zu erkennen. Sie liegen in einer blindsackartigen Abzweigung der holozänen Weseraue, die oberflächlich aus Sand oder Schlick über Kiesen besteht. Nur 275 m von unserer Probenentnahmestelle entfernt wurde 1908 die Tiefbohrung Barbara 1908" niedergebracht (GRUPE 1929a). Die holozänen Ablagerungen sind hier 8 m mächtig. Sie werden bis 20 m Tiefe vom Röt (Oberer Buntsandstein) und bis 389 m vom Mittleren und Unteren Buntsandstein unterlagert.
Die Ursache dieser Hohlformen ist geologisch noch nicht geklärt. Ihrer Form nach erinnern sie stark an Dohnen. GRUPE (1929a) erwähnt sie in seinen Erläuterungen zur Geologischen Karte von Preußen, Blatt Höxter" nicht. Der Nachweis von ausgelaugten Gipslagern an der Röt-Basis, die GRUPE (1929b) für das benachbarte Blatt Holzminden anführt, läßt gleiches für die Entstehung der Grundlosen vermuten. Das unterste Röt ist in ihrem Bereich noch 20 m mächtig. Gleichwohl ist die Gipsauslaugung nicht die einzige denkbare Ursache unserer möglichen Erdfälle. Nach KNAPP (1983) sind
sehr wohl auch Subrosionen von viel tiefer liegenden Zechstein-Salinaren in Betracht zu ziehen.
Der Name haftet diesem Gelände spätestens seit Beginn der Neuzeit an (1467, 1531, 1571). Für den gleichen oder einen unmittelbar daneben liegenden Platz galt auch der Name Kolk: in der Kolcht" (1371), auf der Kolck" (1375), auff der Kolke" (1480, 1593, 1597, sowie in einer undatierten, nach BRÜNING (mdl. Mitt.) um ca. 1350 anzusetzenden Grundstücksskizze (LEESCH 1961, Originaldokument II, 207 in Ziegenhirts Kopialbuch)). Kolk" bezeichnet noch heute im hiesigen Sprachgebrauch ein rundliches stehendes Gewässer, Grundlose" eher einen ausgefüllten Einsturztrichter (vgl. auch POTT 1985a: 60). Die Kiesgrubenarbeiter bezeichnen mit Grundlose" übrigens auch einen bestimmten Unterwasserbagger.
Die Umgebung
Genauere Erkenntnisse über die Bodenbedeckung im weiteren Umfeld der Grundlosen lassen sich erst aus der Zeit gewinnen, seit es zuverlässige topographische Karten gibt. Lediglich die Waldgrenzen zur Weser hin sind, zumindest für den Solling, schon von KRABBE (1603) sehr genau dargestellt worden. Es ist auch zu erkennen, daß sie bis 1759 stellenweise zurückgedrängt worden sind (KLEINAU et al. 1977). Die unverwertbaren Karten von GIGAS vom ehemals corveyischen Gebiet werden von etwa 1600 an bis 1762 immer wieder neu gestochen. Auch die französischen Karten aus den Siebenjährigen und den Napoleonischen Kriegen sagen zu wenig aus. Hingegen malt SIES (auch SIEHS geschrieben) (1704) das Gelände zwischen Godelheim und Höxter überaus genau.
Prozentberechnungen der Bodenbedeckung ermöglicht aber erst die moderne Kartographie, die um 1830 mit dem Urkataster in Preußen und mit den entsprechenden Forstrissen in Braunschweig beginnt. Welch starke Verschiebungen das 19. und 20. Jahrhundert noch brachten, läßt sich aus den nachfolgenden Ausgaben der Meßtischblätter (heute TK 25) ablesen (Abb. 3). Über die Artenzusammensetzung der Wälder und Forsten erfährt man erst spät etwas. MERKEL (1930) zitiert für die Corveyer Besitzungen erstmals für 1564 Eichen, Buchen und anderes Holz". Dabei bleibt es lange Zeit. Etwa 1750 pflanzt Corvey die ersten Fichten, ab 1785 auch Waldkiefern, Edeltannen und Lärchen, sogar auch Edelkastanien, und ab 1800 Schwarzkiefern.
Für den Stadtwald Höxter verzeichnet RODE (1844) folgende Anteile:
Buche, 1-80jährig über die Hälfte
aber unter 20jährig 64,7 %
Eiche, 21-80jährig 1,2 %
Birke, 21-40jährig 1,3 %
Lärche, 1-20jährig 2,5 %
Fichte, 1-40jährig 1,9 %
Kiefer, 1-40jährig 1,4 %
Blößen, schon auf den angegebenen Forstorten
bei SIES (1704) deutlich zu erkennen 25,7 %
"Unland" 1,3 %
Aus der Försterei Boffzen am Sollingfuß schätzt eine Aufstellung aus der gleichen Zeit die Erträge aus Nadelholz zwischen den Jahren 1823 und 1852 auf 1,5 % des Holzeinschlages ein. Bezüglich der Birken ist zu bemerken, daß 1823 auf dem Ziegenberg eine Birkenanpflanzung entstand. Außerdem wurde eine Birkenaussamung für den abzutreibenden Niederwald vorgesehen. Die Birken verschwanden auf dem Muschelkalk aber rasch.
Wälder für Mast, auf denen auf Anordnung der Fürstäbte die Buchen sogar geköpft wurden, um ihre Fruchtbarkeit" zu erhöhen (gemeint ist sicherlich der Blattertrag, vgl. BURRICHTER & POTT 1983, POTT 1990), lagen weiter im Norden des Corveyer Ländchens. Die Höxteraner trieben ihre Schweine gemäß alten Verträgen in den braunschweigischen Solling. Die städtischen Wälder blieben der Weide und Holzung vorbehalten. Die Ziegen ästen seit altersher auf dem Ziegenberg, der erstmals 1365 als "Ceghenberg" erwähnt wird. Im Jahr 1758 waren es 359 Tiere, im Jahr 1803 noch 165 (PREYWISCH 1962). Reste der "Wachholderheide" sind noch heute am Steilhang des Ziegenbergs zu erkennen.
Maßnahmen gegen die Waldzerstörung sind schon früh erkennbar. Mitte des 14. Jahrhunderts wird in einer städtischen Urkunde ein "Vorstenheger" erwähnt, 1371 ein "Hegerweg"
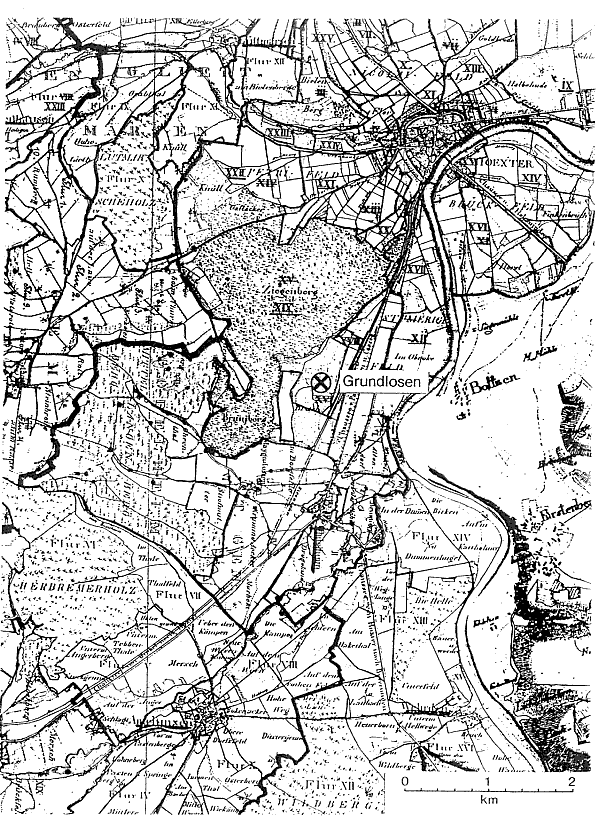
Abb. 3: Die Bodenbedeckung um die Grundlosen um 1830. Die preußischen Urkataster der einzelnen Gemarkungen und die braunschweigischen Forstrisse Boffzen und Derenthal (K 136 u. K 137 des Niedersächsischen Staatsarchivs Wolfenbüttel) wurden auf einheitlichen Maßstab gebracht u. zusammenmontiert. Für diese Arbeit danken wir dem Katasteramt des Kreises Höxter sehr.
(LEESCH 1961). Immer wieder versuchten auch die Äbte als Landesherren die Überweidung, vor allem durch Ziegen, einzuschränken.
Die Schutzmaßnahmen erwiesen sich aber erst unter der kurzen Herrschaft der Oranier um 1803 als einigermaßen wirksam.
Auch über die Landwirtschaft liegen nur zufällige Nachrichten vor. Von den häufigeren Getreidearten wird Roggen um 1350 erwähnt. Besser sind die Überlieferungen von Sonderkulturen im Stummer Feld (LEESCH 1961), das in der Nachbarschaft der Grundlosen liegt. Dort ist in den Jahren 1357, 1365, 1405, 1477 und 1495 die Rede von "Hoppengharden", "Hopfenberg" und "Hopfenhof" am Fuß des Brunsberges. Im Bereich der Grundlosen selbst spricht man 1477, 1484 und 1495 von einem "Wydenbroke", wo die Bürger von Höxter Ruten schneiden. Bei SIES (1704) sind an dieser Stelle zwei helle Gebüsche gut zu erkennen. Weiter entfernt, bei Godelheim, wird um 1500 eine "Petercillienwiese" genannt.
Genauere botanische Nachrichten setzen mit GUTHEIL (1837) ein. Dann sind besonders JÜNGST (vor allem 1852, und da schon mit Hilfe von BECKHAUS) und BECKHAUS selbst (1893) zu nennen. Für den Ziegenberg werden von den drei Autoren somit ausdrücklich Taxus baccata und Sorbus torminalis seit 1837 bestätigt. Heute sind die im Pollendiagramm aufgeführten Baum- und Straucharten alle im Nahbereich vertreten. (Vgl. auch RUNGE 1955, 1972, HAEUPLER 1976). Der nächstgelegene Bestand von Castanea sativa stockt auf dem Steinberg bei Wehrden, 3,5 km südsüdwestlich entfernt. Von den krautigen Pflanzenarten ist nur Plantago maritima für die letzten 150 Jahre nicht angegeben. Centaurea cyanus ist seit etwa 1950 weitgehend verschwunden.
Die Grundlosen
Unmittelbar in und an den Grundlosen wuchsen bis 1963 die Doldenblütler Oenanthe aquatica und Oe. fistulosa. Man findet sie jetzt wenigstens wieder nahebei. Sehr reichhaltig ist hier der Artenbestand der Gattung Carex: C. acutiformis, C. gracilis, C. hirta, C. muricata, C. otrubae, C. riparia, C. rostrata, C. vesicaria und C. vulpina, um nur die hervorstechendsten zu nennen. Von den Gramineen bildete Glyceria maxima am Rand der Großen Grundlose bis 1963 einen schmalen Gürtel um einen solchen von Ranunculus lingua (GUTHEIL 1837, JÜNGST 1852, BECKHAUS 1893). Bis heute hat sich dort der artenreiche Saum des Scirpo-Phragmitetum nicht wieder ausgebildet, nachdem er 1963 durch die flächenhafte Bekämpfung einer um ca. 1950 angelegten Korbweidenkultur mittels Herbiziden zerstört wurde. Dafür hat sich Glyceria maxima großflächig ausgebreitet, auch in Richtung auf die anschließenden Seggenrieder. Sparganium erectum ist gut, Sparganium emersum schwach vertreten. Nach BECKHAUS (1893) hat Typha angustifolia in einigen Exemplaren zwischen Typha latifolia auf dem Bruch neben der Eisenbahn gestanden, scheint aber damals bereits verschwunden gewesen zu sein. Seit 1963 wächst keine Typha-Art mehr in und an den Grundlosen, wohl aber beide Arten an den Kiesgruben, Typha latifolia weit überwiegend.
An Schwimmblatt- und submersen Phanerogamen beherbergten die Grundlosen zwischen 1837 und 1963 ein weit nach Südosten vorgeschobenes Vorkommen von Hottonia palustris. 1976 wurden in der damals separat existierenden und schon längere Zeit trockengefallenen Kleinen Grundlosen noch kleine Landformen von Hottonia beobachtet (vgl. RAUS 1977). Von allen Wasserpflanzen ist heute offenbar nur mehr Lemna minor vorhanden. Die Nordöstlich benachbarte Kiesgrube hingegen hat einen überreichen Bewuchs an Hottonia, Myriophyllum spicatum, Ceratophyllum demersum und Potamogeton crispus, um nur die häufigsten zu nennen.
Das Glycerietum maximae stand meist knietief unter Wasser. Es hatte durch einen Graben bis zum trockenen Herbst 1985 fast ständig Verbindung mit der Kiesgrube. Je nach Wasserstand der Weser fand Wasseraustausch in wechselnder Richtung statt. Große und Kleine Grundlose sind inzwischen zu einem einheitlichen Gewässer zusammengewachsen und sind demgemäß in der Deutschen Grundkarte 1 : 5000 auch so dargestellt.
Palynologische Untersuchungen (Averdieck)
Sondierungen und Bohrungen
Anhaltspunkte für die Entstehungszeit und Entwicklung der Hohlformen selbst vermitteln die pollenanalytisch-paläobotanischen Untersuchungen ihrer Ausfüllung, besonders in Verbindung mit den 14C -Datierungen. Zudem bietet sich die Gelegenheit der vegetationsge-
schichtlichen Forschung innerhalb der Talaue der Weser, reizvoll vor allem angesichts der Vergleichsmöglichkeit mit den Untersuchungen der nahen Gebirgsmoore. Während die Moore, meistens Hochmoore, in den Mittelgebirgen Nordwestdeutschlands schon früh zu palynologischen Studien herangezogen worden sind und ein gutes Bild der Vegetations- und Siedlungsgeschichte geliefert haben (vgl. FIRBAS 1952: 75 - 95, dort weitere Literatur), war die Situation in den Flußtälern ungleich schlechter. Hochmoore konnten hier nicht aufwachsen, und Niedermoore, z. B. aus verlandeten Altwässern, fanden offenbar auch keine Entstehungsbedingungen. So liegen aus dem Weser- und Leinetal nur sehr wenige Pollenanalysen vor, die fast sämtlich an Auelehm und Flußton praktiziert worden sind (NIETSCH 1955, WILLERDING 1960).
Die nächstgelegenen Untersuchungen stammen aus den Sollingmooren und aus dem Bruchwaldmoor im einstigen Weserumlauftal bei Bodenfelde, sämtlich keine 15 km von den Grundlosen entfernt. In etwa doppelter Entfernung liegen die Untersuchungen an Mooren des östlichen Westfalens.
Ein interessanter Bezug ergibt sich zum Ziegenberg, an dessen Fuß die Grundlosen liegen. Dieser zeitweise stark von Siedlungseinflüssen betroffene Muschelkalk-Höhenzug ist durch Vorposten südöstlicher Florenelemente ausgezeichnet (vgl. BUDDE 1951, RAUS 1977, ROHDE 1983). Da es auf dem Ziegenberg selbst keine Moore gibt, galt es zu prüfen, ob Zeugnisse seiner Vegetation und ihrer Veränderungen bis zu den Grundlosen gelangt wären.
Das Gebiet der Grundlosen ist teilweise und sehr unterschiedlich vermoort. Leider ist eine detaillierte geologische Moorkartierung bisher nicht durchgeführt worden. Wir mußten uns, da für das Bohrprogramm nur wenig Zeit zur Verfügung stand, auf nur 20 Pürkhauer-Sondierungen beschränken, um uns eine Übersicht zu verschaffen und die drei Kernbohrungen an den geeignetsten Stellen ansetzen zu können. Die größten Mächtigkeiten von Torf und weichen Sedimenten trafen wir im Norden an. Hier liegen einige der dolinenartigen Teiche sehr nahe beieinander. Nach unseren Pürkhauer-Sondierungen scheint es sich bei diesem Areal um eine gemeinsame größere Einsenkung mit mehreren Restteichen zu handeln. Wie die aufgeführten Bohrprotokolle zeigen, ist die Mächtigkeit der muddigen Oberflächenschichten unabhängig von der Mächtigkeit der unterlagernden organogenen Schichten. Zu diesem Zeitpunkt führte nur der größere Teich der Gruppe eine geringe Wassermenge, so daß Abteufungen in oder nahe dem Zentrum der Teiche leicht möglich waren. Von den Bohrungen sind nachfolgend nur die drei interessantesten protokollarisch aufgeführt.
Pürkhauer-Sondierung nahe dem Zentrum der Hohlform 5:
| 0-215 cm | Mudde, tonig, humos, bis 35 cm sehr weich, mit Schnecken |
| - 240 cm | Ton, grau, mit geröllartigen Mudde-Einschlüssen |
| - 250 cm | Schluff, feinsandig, grau |
| - 270 cm | Schluff wie voriger, rötlich gefleckt |
| - 300 cm | Feinsand, rötlich |
Pürkhauer-Sondierung Zentrum der Hohlform 4:
| 0 - 60 cm | Mudde, tonig, lagenweise mit grobem Pflanzenmaterial |
| - 255 cm | Mudde wechselnder Festigkeit und Farbe, nach unten zunehmend tonig und mit Hölzern |
| - 265 cm | Ton, hellgrau |
| - 375 cm | Torf/Mudde/Ton-Wechsellagerung mit Schnecken und Hölzern |
| - 380 cm | Schluff-Ton, grau |
| - 530 cm | Bruchwaldtorf, holzreich, zeitweilig tonig |
| - 540 cm | Schluff, tonig, mit Holz und Zweigen |
| - 600 cm | Ton, in Schluff übergehend, hellgrau |
Die nachfolgend aufgeführte Schichtfolge entstammt der vollständigsten von insgesamt drei Kernbohrungen, die wir nahe dem Punkt 4 an der Stelle maximaler Gesamtschichtenmächtigkeit auf dem schmalen Steg zwischen den Hohlformen bzw. Teichen 4 und 5 niederbrachten. Für die Kernbergung setzten wir das Klemmkolbengerät nach MERKT und STREIF (1970) ein. Die später zu besprechenden Oberflächenproben für pollenanalytische Vergleichszwecke haben wir acht Jahre später im Juli 1984 anläßlich einer Geländebegehung innerhalb verschiedener Pflanzengesellschaften entnommen.
Kernbohrung Punkt 17, nahe Sondierpunkt 4, zwischen den Hohlformen:
| 0 - 10 cm | Wurzelfilz |
| - 54 cm | Riedtorf mit Equisetum, muddig und tonig, holzreich, mit Mollusken |
| - 62 cm | Ton mit pflanzlichen Einschlüssen |
| - 93 cm | Torf oder Mudde, lagenweise stark tonig, mit wechselndem Holzgehalt |
| - 273 cm | Bruchwaldtorf, holzreich |
| - 525 cm | Bruchwaldtorf wie voriger, z. T. mit großen Ästen, mit zahlreichen geringmächtigen Mudde-, Ton- und Feinsandlagen, besonders um 275, 345, 390 und zwischen 430 und 510 cm. Von 320 - 370 cm auffallend viel Sphagnum |
| - 535 cm | Übergang Bruchwaldtorf/ humoser Ton |
| - 580 cm | Ton, oben humos, nach unten hellgrau werdend, mit Holz-und Torfeinschlüssen |
| - 590 cm | Mudde, tonig, mit Tonschichten, mit Übergang in humosen Ton |
| - 600 cm | Feinsand/Schluff, grau-braun |
| - 603 cm | Bruchwaldtorf, verwürgt |
| - 606 cm | Ton, stark humos |
| - 700 cm | Schluff, tonig, grau, nach unten hin rosa werdend, mit Hölzern und geringmächtigen Ton- und Torflagen |
| - 720 cm | toniger Bruchwaldtorfund Schluff in Wechsellagerung |
| - 735cm | Schluff, stark sandig, rosa, mit Ton- und Stein-Einlagerungen |
| - 740 cm | torfiger Ton |
| - 787 cm | Feinsand, schluffig, rosa, mit einzelnen torfig-humosen Bändern und mit Steinen |
| - 802 cm | Sand bis Kies, rötlich, mit torfigen Einschlüssen |
| - 815cm | Schluff, humos, mit Steinen. Weiterbohrung wegen Steinen nicht möglich. Holozän vermutlich nicht durchteuft. |
Zur Methode
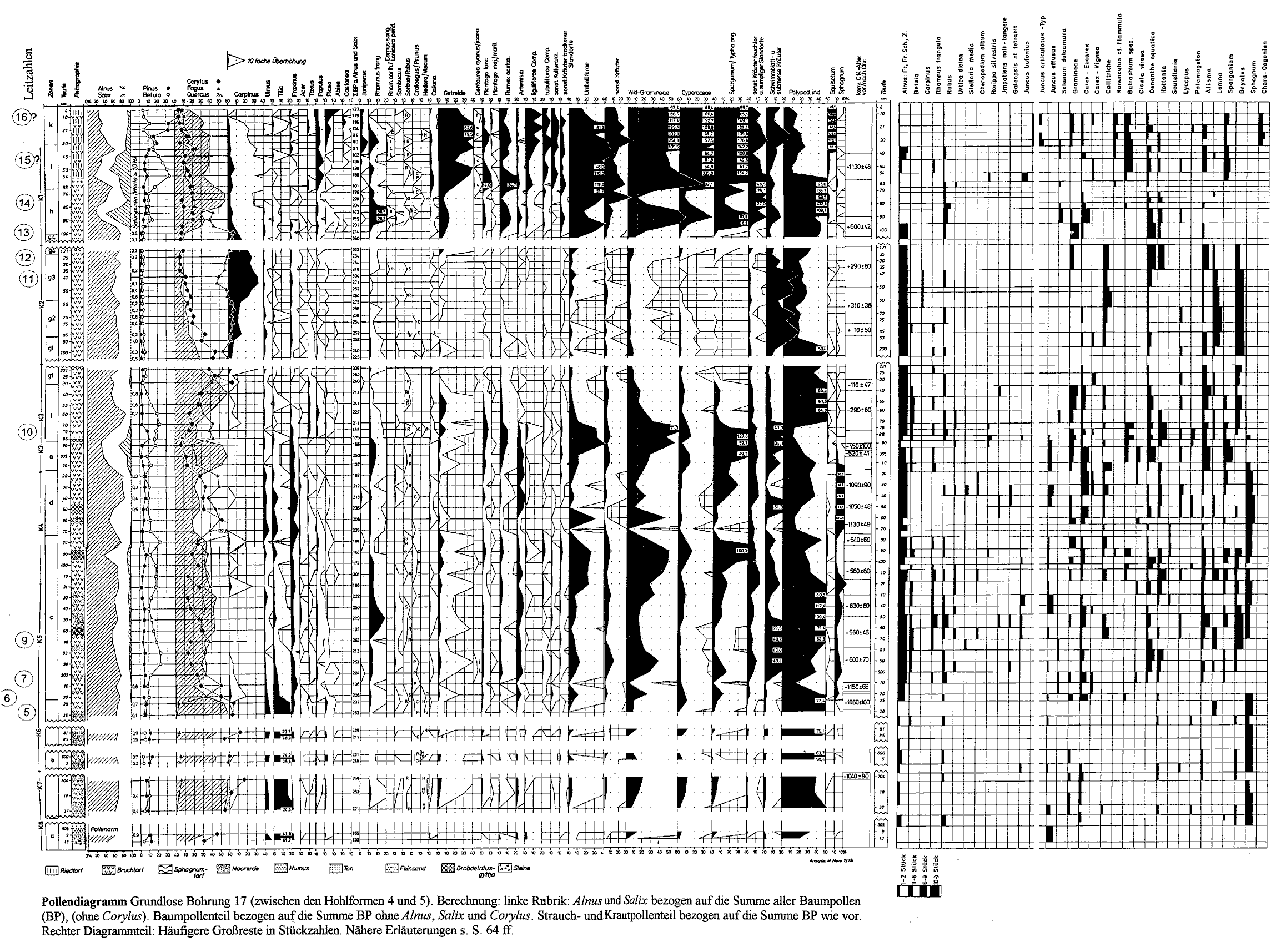
Für die Aufbereitung zur Pollenanalyse wurde die Kalilauge-Acetolyse-Methode nach ERDTMAN (1954) angewandt, zur Makrofossil-Analyse schwache Salpetersäure. Die Ergebnisse der Pollenanalysen sind im Pollendiagramm und in Tabelle l wiedergegeben. Wegen der allgemein starken Überrepräsentanz von Alnus (bis fast 90 % der Baumpollen) und stellenweise auch von Salix wurden für die Berechnung zwei verschiedene Grundsummen benutzt. Alnus und Salix sind auf die Gesamtpollensumme bezogen und in der ersten Rubrik so dargestellt, daß die freie Fläche die restlichen Baumpollen bezeichnet. Sämtliche anderen Frequenzen beziehen sich auf die Summe dieser restlichen Baumpollen. Auch die rechts von der Bezugssumme stehenden Strauch- und Krautpollen sind auf die Baumpollensumme ohne Alnus und Salix bezogen. Die hieraus resultierende Überhöhung gegenüber der konventionellen Berechnungsweise, wie z. B. bei den Solling-Diagrammen, ist zu beachten. Andererseits ist der Vergleich mit ähnlich berechneten Diagrammen (WILLERDING 1960, LESEMANN 1968, POTT 1982, 1985a und b) erleichtert.
In dem Diagramm sind außer den Gehölzen nur die für unsere Fragestellung bedeutungsvollsten Kulturanzeiger und einige besonders häufig vertretene Taxa aufgeführt. Der Rest ist in Anlehnung an die vermuteten Biotope in Summenkurven zusammengefaßt, einzeln aber in der Tab. l verzeichnet. Die häufigsten Makrofossilfunde, meistens Früchte oder Samen, sind in dem Stückzahl-Diagramm dem Pollendiagramm angefügt, die selteneren in Tab. 2 aufgeführt.
Zum Pollendiagramm
Das Diagramm zeigt auf den ersten Blick die auffällige Zweigliederung, wie sie auch in den Solling-Diagrammen (HESMER 1928, KNÖRZER 1949, STECKHAN 1961, SCHNEEKLOTH 1967) zum Ausdruck kommt.
{kind=link}
In erster Linie nach den Baumpollen (ohne Berücksichtigung von Alnus und Salix), aber auch nach den Kulturanzeigern ist unser Diagramm in zehn charakteristische Abschnitte unterteilt, die auch die Parallelisierung mit aus der Umgebung stammenden Diagrammen erleichtern.
Die besondere Problematik, die durch die 14C-Daten herangetragen wird, bleibt bei der nachfolgenden kurzen Charakterisierung unberücksichtigt.
Abschnitt a (813 - 805 cm): Maximalwerte von Tilia, Fagus noch fehlend. Kulturanzeiger vereinzelt.
Abschnitt b (737 - 525 cm): Tilia, Corylus, Quercus und, weniger ausgeprägt, auch Ulmus mit hohen Werten, Fagus niedrig, aber regelmäßig, Carpinus sporadisch. Kulturanzeiger inkl. Getreide durchgehend.
Abschnitt c (520 - 380 cm): Fagus-Steilanstieg mit erstem Gipfel, dann Fagus und Quercus ziemlich gleich hoch, Tilia, Ulmus und Corylus rasch abfallend, Fraxinus etwas stärker, Carpinus weiterhin sporadisch. Kulturanzeiger durchgehend ziemlich hoch.
Abschnitt d (370 - 320 cm): Fagus dominierend über Quercus, übrige Eichenmischwald-(EMW-)Glieder etwas reichlicher, Corylus mit Maximum. Kulturanzeiger erniedrigt, besonders Getreide zeitweilig aussetzend.
Abschnitt e (310 - 290 cm): Kurzes Quercus-Maximum, Fagus-Minimum, Corylus-Abfall. Wiederanstieg der Kulturanzeiger.
Abschnitt f (285 - 240 cm): Wechselnde Fagus/Quercus-Dominanz, Beginn der geschlossenen Carpinus-Kurve. Kulturanzeiger-Maximum.
Abschnitt g (230 - 100 cm): Fagus-Dominanz und absolutes Maximum, Carpinus-Maximalwerte, Quercus beständig fallend, Corylus nach letztem Gipfel auf Minimalwerte sinkend. Kulturanzeiger lückenhaft. Dieser Abschnitt ist zu untergliedern:
g1 (230 - 193 cm): Carpinus langsam ansteigend, Corylus letztes Maximum.
g2 (185 - 160 cm): Carpinus-Stufe über 10 %, Fagus-Maximum. Absolutes Kulturanzeiger-Minimum.
g3 (155 - 125 cm): Carpinus-Maximum, leichte Fagus-Depression, Quercus- und Corylus-Minimum.
g4 (121 - 100 cm): Letztes Fagus -Maximum, Carpinus abfallend. Kulturanzeiger-Anstieg.
Abschnitt h (95 - 70 cm): Quercus-Maximum, Fagus-Abfall, Carpinus niedrig, Alnus-Depression, Salix-Maximum zugleich mit Rhamnus frangula-Maximum. Kulturanzeiger noch mäßig, im oberen Teil schwach rückläufig, Secale-Auftreten.
Abschnitt i (63 - 35 cm): Quercus dominierend, Betula-Maximum, Fagus und Corylus niedrig, Populus- (und Juniperus-)Maxima. Starker Kulturanzeiger-Anstieg (mit nachfolgendem Einschnitt bei 45 cm), Centaurea cyanus ziemlich regelmäßig.
Abschnitt k (30 - 4 cm): Pinus-Anstieg zur Dominanz, Picea-Anstieg, Juniperus-Maximum. Getreidekurve nach Maximum rückläufig.
Die waldgeschichtliche Gliederung nach den mitteleuropäischen (FIRBAS 1949 = Fi) oder nordwestdeutschen Pollenzonen (OVERBECK 1975 = Ov) führen wir am besten über die Parallelisierung mit den Solling-Hochmooren durch, wobei die Zonierung nach FIRBAS (1952: 78 f.) für das Mecklenbruch und deren Verknüpfung von OVERBECK (1975: 529) zugrunde gelegt werden. Für die Charakterisierung und Konnektierung der Solling-Diagramme haben sich die von STECKHAN (1961: 540 f.) aufgestellten (und hier auf die Solling-Moore zugeschnittenen) Leithorizonte bewährt, so daß sie im Zuge der vergleichenden Betrachtung auch für das Grundlosen-Diagramm heranzuziehen sind (als kreisumrandete Ziffern im Diagramm eingetragen). Die infrage kommenden Leithorizonte sind wie folgt definiert (in etwas abgekürzter Fassung):
b) Eichenmischwaldzeit
4 Schwacher Abfall der Quercus-Kurve, Beginn der nahezu geschlossenen Carpinus-Kurve und des letzten Haselgipfels, Ende der Periode mit höheren Plantago-Werten.
5 Beginn des endgültigen Eichenrückganges, schwache Zunahme der Buchenwerte, Endphase des letzten Corylus-Gipfels.
c) Eichenmischwald-Buchenzeit
6 Rationelle Buchengrenze (starker Anstieg der Kurve).
7 Kreuzung EMW x Fagus und Alnus x Fagus.
8 Beginn der Periode mit Getreidepollen und höheren Plantago-Werten (nicht in Grundlose zu übernehmen).
9 Erster Buchengipfel.
d) Buchenzeit
10 Beginn der endgültig geschlossenen Carpinus-Kurve, Buchenminimum, Birkengipfel.
11 Buchenminimum (im Mecklenbruch über mehrere Spektren ausgedehnt), erhöhte Birken- und Erlenwerte, Anstieg der Carpinus-Kurve.
12 Beginn der geschlossenen Getreidekurve, schwacher Rückgang der Hainbuche, schwache Zunahme von Eiche und Hasel (nach KNÖRZER 1949 im Mecklenbruch Beginn des Buchenrückganges bei α).
13 Starker Buchenabfall (besonders im Torfmoor), starker Anstieg der Kurven von Birke, Getreide, Plantago lanceolata, Wildgräsern und Siedlungsbegleitern - entspricht etwa β im Mecklenbruch.
e) Zeit der Waldnutzung
14 Eichengipfel, Rückgang von Carpinus (endgültig) und Corylus, hohe Getreide-, Wildgras- und "Unkräuter"-Werte, die Pinus-Werte steigen auf etwa 10 % -entspricht ungefähr 8 im Mecklenbruch.
15 Kleiner Buchengipfel (in beiden Mooren), entspricht ε im Mecklenbruch.
f) Zeit der geregelten Forstwirtschaft
16 Picea-Maximum, Getreidemaximum, Eichenrückgang.
Im Diagramm der Grundlosen gehört der Abschnitt a vielleicht noch dem Ausklang des Atlantikums, der Zone VIII Ov (VII Fi) an. Die bereits nachweisbaren Kulturanzeiger brauchen nach den siedlungsgeschichtlichen Ergebnissen von WILLERDING (1965, 1971) und POTT (1982) nicht zu stören.
Abschnitt b gehört in die subboreale Zone IX Ov (Villa Fi), wie aus der beherrschenden Rolle des EMW bei gleichzeitig hoher Corylys-Vertretung zweifelsfrei hervorgeht. Wahrscheinlich liegen nur kurzzeitige Ausschnitte vor, wie später noch zu diskutieren ist. Bemerkenswert sind die hohen Tilia-Werte (allerdings durch Alnus-Herausnahme überhöht). Derartige Werte sind von Bodenproben her bekannt und können, trotz Begünstigung durch Zersetzungsauslese, als Ausdruck starker bis dominierender Lindenbeteiligung an manchen EMW-Formen gelten (vgl. HAVINGA 1962, ANDERSEN 1973). Dem Bewuchs trockener Standorte entspricht die Pollenzusammensetzung örtlicher Substrate (wie z. B. Oberböden) sicherlich besser als der von Seesedimenten und Mooren, selbst wenn nahe gelegen, mit ihrem andersgearteten Um- und Eigenbewuchs. Ob im vorliegenden Fall die Lindenpollen aus einer Hartholzaue stammen, wie es LESEMANN (1968) für das Auenprofil bei Bodenfelde vermutet, oder ob sie aus höheren Lagen stammen, läßt sich für unsere Spektrenfolge ohne weitere Anhaltspunkte nicht entscheiden. Die basalen Proben mögen für erstere Deutung sprechen, die folgenden eher für die zweite, denn der Lindenpollen erreicht bei gleicher Berechnungsweise auch im Torfmoor auf dem Solling bis zu 36 % im Boden unter dem Moor und 28 % im Älteren Hochmoortorf. Hier wie dort verhält sich die Linden- wie die Eichenkurve gegenläufig zur Buchenkurve, während Esche und Erle unbeeinflußt bleiben.
Der Auenwald der Umgebung hat in der Späten und auch in der Älteren Nachwärmezeit offenbar mehr den Charakter von eschenreichen Erlenauenwäldern getragen, wie sie WILLERDING (1960) für die Göttinger Leineauen folgert. Die von STECKHAN (l961) genannten Leithorizonte 4 und 5 kämen für diese Zone in Betracht, sind aber wegen der Unterbrechungen in diesem Diagrammabschnitt nicht näher lokalisierbar.
Abschnitt c schließt die hier sehr kurze Übergangszone vom Subboreal zum Subatlantikum und den älteren Teil des Subatlantikums ein (X und XI Ov bezw. VIIIb und IX Fi). Die fast abrupt einsetzenden Veränderungen, die mit den Leithorizonten 6 (Buchenanstieg) und 7 (Kreuzung der Buchen- und Eichenkurven) zu belegen wären, lassen einen Hiatus vermuten. Außer einigen Kurvensprüngen deuten auch die 14C-Datierungen in diese Richtung. Dennoch müssen sich diese Wandlungen sehr rasch vollzogen haben, denn sie sind, nicht minder ausgeprägt, auch in den Solling-
Diagrammen zu finden. Für das Torfwachstum der Solling-Hochmoore aber wäre kaum eine Unterbrechung oder Störung zu postulieren, wie man solche vielleicht bei einem Erdfall erwarten könnte.
Der Vergleich der 14C-Daten für diesen Horizont zwischen Solling und Grundlosen ergibt überraschenderweise ein zwischen 300 und 500 Jahre höheres Alter bei letzterer, wobei hier die Datierung sogar die jüngere Stelle des Leithorizontes 9 betrifft. Der ausgedehnte Abschnitt c enthält nach anfänglich kurzem Fagus-Maximum recht ausgeglichene hohe Fagus- und Betula-Frequenzen, letztere vielleicht im Einklang mit den deutlichen Siedlungsaktivitäten. Im Gegensatz zu den Solling-Mooren ist die mit dem nächsthöheren Leithorizont 10 (Beginn geschlossener Carpinus-Kurve) zu bezeichnende Veränderung in einem sehr viel größeren Abstand zu finden. Dennoch ist der Abschnitt nicht ohne Dynamik, angezeigt vor allem durch lokale Elemente.
Abschnitt d, insbesondere durch Fagus- und Corylus-Maxima bei höheren Ulmus-, Tilia- und Fraxinus-Werten ausgezeichnet, findet in den Solling-Diagrammen keine Parallele. Auffällig ist die Lücke in der Getreidekurve, die von den übrigen Kulturbegleitern zwar auch angedeutet wird, aber verschoben und ungleichmäßig. Da Carpinus fast völlig fehlt, muß auch dieser wie der folgende kurze, stark von menschlichen Eingriffen geprägte Abschnitt e und der untere Teil des nächsten Abschnitts f dem frühen Subatlantikum zugerechnet werden.
Im Abschnitt f findet sich mit dem Einsetzen der Carpinus-Kurve der nicht exakt lokalisierbare Leithorizont 10. Im weiteren Diagrammverlauf lassen sich die Leithorizonte von STECKHAN (1961) ohne Schwierigkeit ansetzen, jedoch ist die Übereinstimmung der Merkmale nicht in jedem Fall eindeutig, insbesondere nicht bei den mit Siedlungsanzeichen begründeten. Abschnitt g wird von Fagus und Carpinus charakterisiert. Vom obersten Unterabschnitt g4 abgesehen, sinken die Anzeichen menschlicher Aktivität auf ein Minimum, anfänglich gefolgt von kräftiger Haselausbreitung in g1.
Der Leithorizont 11 dürfte bei dem Carpinus-Maximum und dem kleinen Fagus-Minimum in g3, der Leithorizont 12 beim Wiedereinsetzen der Kulturanzeiger an der Grenze g3/g4 anzubringen sein.
Während sich die zunehmende Siedlungsaktivität bei den Baumpollen zunächst lediglich in einer Beeinträchtigung von Carpinus widerspiegelt, leitet der Abschnitt h, an dessen Beginn der Leithorizont 13 (Buchenabfall, Birken- und Siedlungszeigeranstieg) gehört, zum Jung-Subatlantikum, Zone XII Ov bzw. X Fi über. Starke Veränderungen im Landschaftsbild sowohl der Flußaue als auch der Hochlagen kommen eindrucksvoll heraus, nämlich die allgemeine Walddezimierung und Bodendegradierung.
Von den obersten Leithorizonten mag 14 noch mit der Plazierung am Fagus-Abfall und Quercus-Maximum inmitten des Abschnitts h zu vereinbaren sein.
Die Leithorizonte 15 und 16 sind angesichts der lokalen Abweichungen kaum mehr auf das Grundlosen-Diagramm übertragbar. Die Zonen-Untergliederung Xa/Xb Fi würde sich etwa mit der Grenze zwischen den Abschnitten i und k zur Deckung bringen lassen.
Rein pollenanalytisch-vegetationsgeschichtlich stehen die hier betrachteten Diagramme in guter Übereinstimmung miteinander. Vergleicht man die absoluten Daten, die korrelierbaren Horizonten zuzuordnen sind, so ergeben sich jedoch erhebliche Differenzen. Für die Solling-Moore stehen die 17 von SCHNEEKLOTH (1967) vorgelegten und auf die Leithorizonte bezogenen 14C-Daten zur Verfügung.
Am einfachsten erscheint der Vergleich bei dem markantesten Horizont, dem Fagus-Steilanstieg, an dem im Grundlosen-Profil die Leithorizonte 5 und 6 nahe beeinander liegen und ihre 14C-Daten den Zeitraum von 1700 bis 1100 v. Chr. umspannen. Im Solling belaufen sich die Daten für die wesentlich größere Profilstrecke auf die Spanne von 1700 bis 800 v. Chr.. Dabei ist im Solling aber erst der beginnende, in der Grundlosen bereits der abgeschlossene Fagus-Anstieg datiert, so daß dieser bedeutende Vegetationswandel im Solling erheblich später eingetreten sein müßte als in der nächsten Grundlosen-Umgebung.
Die nächstfolgenden Daten liegen zwischen den Leithorizonten 7 und 8, die in der Grundlosen kaum auszumachen sind und etwa mit dem Leithorizont 6 zusammenfallen. Jedenfalls liegen sie im Beginn des Fagus-Maximums und noch vor Beginn der geschlossenen Carpinus-Kurve. Bei dieser allerdings recht
variablen Position besteht mit 600 v. Chr. eine relativ gute Übereinstimmung.
Die Leithorizonte 9 und 10 flankieren Daten um 100 v. Chr. Für eine Verknüpfung kommt nur der Leithorizont 10 infrage, der mit Carpinus-Kurvenschluß, kleinem Fagus-Minimum und Betula-Gipfel eine Gruppe auseinanderliegender und z. T. offensichtlich sehr lokaler Merkmale umfaßt. Für den Beginn der geschlossenen Carpinus-Kurve wurde im Grundlosen-Profil die Spanne zwischen 300 und 100 v. Chr. ermittelt, so daß auch für diesen Teil Übereinstimmung besteht.
Letztlich ist noch der Leithorizont 12 am Beginn des Carpinus-Abfalls vergleichbar. Die Divergenz der Daten "um 700 n. Chr." für den Solling und "zwischen 300 und 600 n. Chr." für die Grundlosen könnte bedeuten, daß die hier reflektierten menschlichen Eingriffe nicht gleichzeitig stattgefunden haben.
Bei dieser Betrachtung sollte allerdings nicht übersehen werden, daß die Daten an unterschiedlichen Torfarten gewonnen worden sind, die erstens für Vergleiche problematisch sind, zweitens ungleiche Voraussetzungen für richtige Datierungen bieten.
Diskussion der 14C-Datierungen
Bei der vegetations- und siedlungsgeschichtlichen Auswertung des Pollendiagramms ist der Besonderheit, die ein Erdfall darstellt, Rechnung zu tragen (vgl. AVERDIECK 1986). Die Absenkung kann einmalig, kontinuierlich oder periodisch erfolgt sein, so daß die Auswirkungen auf die Füllung unterschiedlich sein müssen. Die petrographische Betrachtung der Torfe und Sedimente gibt Hinweise, daß wir mit der letztgenannten Möglichkeit zu rechnen haben. Das gilt insbesondere für den unteren Profilteil mit dem Wechsel von Torf- und Sand- oder Muddelagen. Im oberen Profilteil hingegen sind nur geringfügige Wandlungen konstatierbar, weshalb man für diesen auf gleichmäßige Absenkung schließen könnte. Den gleichen Eindruck vermitteln hier auch einige durchlaufende Makrofossilfunde (Oenanthe, Alisma). Dennoch gehen deutliche Wandlungen aus den wechselnden Dominanzen der lokalen Pollenflora hervor. In dem gemeinhin als Bruchwaldtorf angesprochenen Substrat heben sich nassere Phasen mit Wasserpflanzen gegenüber Verlandungsphasen mit Ufervegetation heraus.
Offensichtlich ist die Sedimentierung oder das Torfwachstum nicht gleichmäßig verlaufen. Den Umfang solcher Schwankungen zu ermitteln, dient schon ein Vergleich der Pollenkurven mit solchen von weniger abhängig gewachsenen Profilen, wie sie Hochmoore bieten. Unsere diesbezügliche Heranziehung der Leithorizonte von STECKHAN (1961) konnte zwar die Ansprüche an die erforderliche Genauigkeit kaum befriedigen, deutete aber schon einige nicht überbrückbare Diskrepanzen an. Ein besseres Ergebnis versprachen wir uns von absoluten Daten. Darum haben wir eine verhältnismäßig große Zahl von 14C- Datierungen über das Profil gelegt, teilweise in engen Abständen (vgl. WILLKOMM, S. 85 in diesem Heft). Die anfänglich viel geringere Anzahl galt den bedeutsamen Diagrammhorizonten.
Während im Profilteil oberhalb 3 m Tiefe die Datenfolge normal verlief, gaben in den unteren vier Metern merkwürdige Erscheinungen Anlaß, auf Störungen zu schließen. Mehrere Zwischenproben erhöhten die Sicherheit der Datenskala. Wie später näher auseinandergesetzt wird, überlagern sich offenbar invers zwei Abschnitte, jeweils mit einer Folge praktisch gleichbleibender Daten. Die obere Folge pendelt um 1100 v. Chr. und nimmt den stratigraphischen Abschnitt d ein, während die untere mit 5 Daten um 600 v. Chr. auf 130 cm Profillänge fast den gesamten Abschnitt c beansprucht. Palynologisch betrachtet, bietet sich innerhalb dieser beiden Abschnitte eine gewisse Uniformität, die aber nicht unbedingt ein Argument für Gleichaltrigkeit abgibt, da so etwas auch in anderen Pollenzonen üblich ist. Von völliger Gleichzeitigkeit kann im übrigen keine Rede sein, denn die sinnvollen Abfolgen der lokalen Nichtbaumpollen sprechen gegen die spontane Ablagerung eines Gemisches von allochthonem Material.
Leichter ist im Diagramm die Inversion der Abschnitte c und d zu interpretieren. So entsprechen einander die Baumpollenfrequenzen des Abschnittes d und des Beginns von c (bei 510 cm). Auch einige auffällige Erscheinungen im Abschnitt d, z. B. das Corylus-Maximum und das Zurücktreten der Kulturanzeiger, ließen sich durchaus am Übergang der
Abschnitte b/c unterbringen. Über diese Ähnlichkeiten hinaus gibt es aber für die Zusammengehörigkeit der beiden Horizonte keine zwingenden Indizien. Die lokale Nichtbaumpollenflora zeigt Eigenentwicklungen, die in den beiden Komplexen unterschiedlich verlaufen und auch nicht auf Störungen hinweisen.
Auch aus dem Substrat selbst läßt sich keine Erklärung ableiten. Der Bruchwaldtorf führt unterhalb 3 m zwar zunehmend schmale Muddeeinlagen, hat aber, nach den lokalen Makro- und Mikroresten zu urteilen, stets amphibischen Ursprung gehabt. Allein schon die Hochwässer der Weser dürften für mehrfache kräftige Wasserspiegelschwankungen gesorgt haben. Diese und mögliche Untergrundbewegungen können Sedimentrutschungen verursacht haben, so daß älteres Material vom Rande auf jüngeres im tiefer gelegenen Zentrum gelangte. Umgekehrt gibt es auch Beispiele für Unterlagerung älterer Schichten durch jüngere infolge Aufschwimmens oder Unterkehlung (vgl. die "klassische" Situation im holsteinischen Duvenseer Moor bei SCHÜTRUMPF 1981). Die genauere Abklärung setzt aber sehr viel umfangreichere Grabungs- und Bohrarbeiten voraus als wir sie leisten konnten. Die vielleicht naheliegende Vermutung einer Bohrkernverwechslung, für die unser Bohrverfahren ohnehin keinen Anlaß bot, entfällt schon deshalb, weil sich die Veränderungen innerhalb der Kerne 4 und 5 abspielen (die Kernnummern sind am linken Rand des Diagramms angegeben).
Da, wie auch immer, die absoluten Daten kaum einen Zweifel lassen, wird man bei weiterer Diagramminterpretation den Abschnitt d besser unberücksichtigt lassen und vom vorhergehenden Abschnitt c annehmen, daß er in einem sehr kurzen Zeitraum zur Ablagerung kam. Was die untersten Daten anbelangt, so wurde auf die Möglichkeit eines Hiatus zwischen den Abschnitten b und c bereits hingewiesen. Für das geringe Alter der Teufe um 704 cm (l 100 v. Chr.) kann nach den diversen gleichalten, aber darüber liegenden und Fagus-beherrschten Proben nur behauptet werden, daß es in Wahrheit höher sein muß.
Siedlungsgeschichtliche Aspekte
Im Pollendiagramm nehmen die Kulturanzeiger einen gesonderten Raum ein. Wenigstens zwei Perioden menschlicher Aktivität heben sich heraus. Daneben künden manche Veränderungen in den Rubriken der Gehölzpollen von Einwirkungen durch Mensch oder Haustier. Leider setzt trotz der großen Gesamtmächtigkeit die durchlaufende Sedimentation zu spät ein, um zu der Frage jungsteinzeitlicher Talauenbesiedlung (WILLERDING 1965, POTT 1982) oder zur Frage der Zugehörigkeit zu einer der westfälischen Grundtypen (BURRICHTER 1970, 1976, POTT 1982, 1984, 1985b) beitragen zu können.
Das Profil zeigt von Anbeginn Kulturspuren, die sich in allen analysierbaren Schichten bis kurz vor der Buchenzeit fortsetzen. Am bronzezeitlichen Alter des Abschnitts b besteht kein Zweifel, für a ist die Datierung unsicher. Spätbronze- bis früheisenzeitlich sind auch die kontinuierlichen, infolge der Berechnungsbasis aber etwas überhöhten und sprunghaften Getreide- und Unkrautpollenkurven der Abschnitte c und e. (Die im Abschnitt d ersichtliche Rückläufigkeit, insbesondere der Getreidekurve, bleibt wegen der möglichen Inversion hier außerhalb der Betrachtung.)
Nach einem kurzen Getreidemaximum im Abschnitt f am Ende des letzten Jahrtausends v. Chr. setzt eine auffällige Rezession während des Abschnitts g ein. Dem Pollendiagramm nach ist es nicht auszuschließen, daß es in der Umgebung zeitweise zu völliger Siedlungsunterbrechung gekommen ist, etwa im Abschnitt g2, doch ist Siedlungsleere während der ersten nachchristlichen Jahrhunderte archäologisch unbegründet (STEPHAN 1978). Zwischen 300 und 600 n. Chr., archäologisch fundiert kaum vor 600, beginnt mit Abschnitt g4 verstärkte Siedlungstätigkeit.
Bald darauf ist erstmals Roggen nachweisbar (Abschnitt h). Ein kurzer Rückschlag könnte im oberen Teil des Abschnitts h angedeutet sein, mit einer Zeitstellung noch vor 1100 n. Chr.. Ab hier beginnen die Steilanstiege der Kulturanzeiger (Abschnitt i), zusammenfallend mit dem Auftreten von Centaurea cyanus. Da noch vor dem 14C-Datum von 1130 n. Chr. gelegen, könnte sich hiermit die Expansion des 9. Jahrhunderts dokumentieren (STEPHAN 1978). Um das genannte Datum herum ist es vielleicht erneut zu einer kurzen Depression gekommen, wofür jedenfalls die Werte der direkten Kulturanzeiger sprechen.
Bezüglich der Beeinträchtigung der natürlichen Vegetation gibt die Parallelisierung der Kurvenschwankungen der Kulturanzeiger mit denen der Bäume und Sträucher einige Aufschlüsse. Spätestens um 1100 n. Chr. dürfte die weitgehende Entwaldung der nächsten Umgebung stattgefunden haben und die Grundlage für die Trockengesellschaften auf dem Ziegenberg gelegt worden sein (BUDDE 1951). Calluna und Juniperus gewinnen zusammen mit Betula und Populus an Bedeutung bei gleichzeitiger Dezimierung von Fagus und Quercus. Tiefgreifende Veränderungen müssen sich auch in der Talaue abgespielt haben, wie die unvermittelten Dominanzwechsel zwischen Alnus und Salix anzeigen. Das plötzliche Hervortreten mancher Pollenkurven (z. B. von Betula, Populus, Rhamnus frangula) mag sich auf Eingriffe sowohl im Tal als auch auf den Höhen beziehen. Während ganz allgemein Quercus von der Siedlungsaktivität unterschiedlich berührt wird, stehen Fagus und Carpinus ganz klar im Gegensatz dazu. Tilia und Ulmus reagieren in den unteren Abschnitten vor allem auf die Fagus-Ausbreitung. Zusammen mit Fraxinus zeigen sie aber auch Reaktionen auf die abgrenzbaren Siedlungsanzeichen. Die verschiedenartigen Bewirtschaftungsweisen der Wälder in Nordwestdeutschland und ihre Auswirkung auf die Folgevegetation hat POTT (1990, dort weitere Literatur) zusammengestellt. In einleuchtender Weise sind von ihm auch die Nachweise in Pollendiagrammen geführt worden (POTT 1982, 1984, 1985a). Im Hinblick auf den Ziegenberg wäre das vorgeschichtliche Auftreten von Taxus von besonderem Interesse (vgl. WILLERDING 1968). Dem Diagramm zufolge hat der Baum seit der Bronzezeit hier aber keine größere Rolle im Waldbild gespielt als im norddeutschen Tiefland (vgl. AVERDIECK 1971). Seine Ausbreitung bezw. Pollenstreu hat offenbar mehr unter der Buche als unter dem Menschen gelitten.
Im Abschnitt k treten mit Pinus, Picea und Castanea bereits die teilweise seit dem 18. Jahrhundert historisch belegten Pflanzungen in Erscheinung. Über die Verknüpfungsmöglichkeit der Kulturphasen zwischen Grundlosen, Solling und den westlich angrenzenden westfälischen Gebirgen vermittelt die Übersicht in Tab. 3.
Die Oberflächenproben
Um einen Anhalt zu gewinnen, in welchem Verhältnis die obersten Spektren des Profils zur gegenwärtigen Umgebungsvegetation stehen, haben wir aus verschiedenen Biotopen Oberflächenproben genommen und auf gleiche Weise analysiert wie die fossilen Proben. Dabei war es unvermeidbar, verschiedene Substrate zu benutzen.
Probe 1 stammt aus Oberflächenschlamm eines breiten Grabens im Erlengehölz.
Probe 2 war ein kleines Moospolster, bestehend aus Eurhynchium stokesii
(TURN.) B.S.G. und Brachythecium rivulare (BRUCH.) B.S.G. von einem
kleinen Acker im Jungfichtenwald.
Probe 3 entstammt ebenfalls einem Moospolster aus Mnium undulatum HEDW.
und Rhytiadelphus squarrosus ( HEDW.) WARNST. aus einem Parkgelände mit
Rasen.
Proben 4 und 5 wurden aus dem Teich 8 aus dem flutenden Moos
Leptodictyum riparium WARNST, bzw. aus flutender Callitriche gewonnen
(sämtliche Moose det. Dr. KLINGER, Oldenburg).
Wie Tab 4 deutlich macht, kommt die Probe 3 dem oberen Diagrammbild am nächsten. Die Rezentspektren demonstrieren eindringlich ihre Abhängigkeit vom örtlichen Bewuchs. Das noch erfaßte Moorstadium im Pollendiagramm wird in mancher Beziehung der benachbarten heutigen Parklandschaft geähnelt haben. Der Pollenflug vom nahen Ziegenberg macht sich in der Probe vom baumlosen Acker am stärksten bemerkbar. Sie enthält auch die höchsten Nichtbaumpollenwerte. Den ziemlich niedrigen Kulturanzeigerwerten in den Rezentspektren nach zu schließen, ist der Ackerbau in unmittelbarer Nähe der Grundlosen früher verbreiteter gewesen als heute. Diese Tendenz ist auch aus der Getreidepollenkurve des Diagramms abzuleiten. Die Neubewaldung des gesamten Geländes dokumentiert sich durch die relativ niedrigen Nichtbaumpollenwerte gegenüber denen des oberen Diagrammteils.
Der lokale Bewuchs
Außer den auf die Umgebung gerichteten geo- und siedlungsbotanischen Forschungen interessiert auch die Geschichte der Hohlform und ihrer Ablagerungen. Die einstigen örtlichen Pflanzengesellschaften, wie sie sich aus sicher bestimmten Pollenfunden und, besser noch, aus den Makrofossilbestimmungen ergeben, sind nach RUNGE (1973) unschwer den Röhrichten und Großseggenriedern (Phragmitetea Tx. et PREISING 1942) zuzuordnen. Offenbar wurden die Flächen neben- oder nacheinander vom Teichröhricht (Scirpo-Phragmitetum W. KOCH 1926),
Wasserschwaden-Röhricht (Glycerietum maximae HUECK 1931) und vom Wasserfenchel-Kressensumpf ( Oenantho-Rorippetum amphibiae LOHMEYER 1950) eingenommen. Zeitweilig bahnte sich der Übergang in ein Weiden-Faulbaum-Gebüsch (Frangulo-Salicetum cinereae MALCUIT 1929) an, während die versumpfte Umgebung bereits von einem Alnion glutinosae MALCUIT 1929 beherrscht war.
An der Besiedlung der offenen Flächen waren Unterwasser-Laichkraut-Gesellschaften (Potamogetonion W. KOCH 1926) sowie Schwimmblatt-Gesellschaften ( Nymphaeion OBERDORFER 1957) beteiligt. Die meisten Gesellschaften weisen auf flache Tümpel mit guter Nährstoffversorgung hin, in die nur stellen- oder zeitweise (Abschnitt d) mesotrophe Verhältnisse einbrachen.
Mit den untersten erfaßten Kiesen und Sanden ist die Holozänbasis von der Bohrung offenbar nicht erreicht worden. Die organogene Ausfüllung macht einen bis in die Gegenwart anhaltenden erstaunlich gleichartigen Eindruck, insbesondere nach den Makro-Funden. Die Anzeichen für nasse Bedingungen lassen Zweifel an der Richtigkeit der Bezeichnung "Bruchwaldtorf" für das holzreiche Bohrgut offen. Obschon auch zahlreiche Funde auf Auenwald verweisen, wäre vielleicht die Ablagerung besser als Grobdetritus-Gyttja, Schwemmtorf oder als Mischung solcher Mudden mit Riedtorf zu bezeichnen. Der umgebende Wald hat sich offenbar mehr durch Hineingefallenes als durch Bewuchs im Sediment oder Torf manifestiert. Jedenfalls sind ständig hoher Grundwasserstand und Überflutungen als sicher anzunehmen. Die erwähnten gleichbleibenden 14C-Daten künden von zeitweilig raschem Torf- oder Sedimentwachstum.
Alle diese Fakten zusammen mit den eingelagerten Sandschichten und den möglichen Sedimentrutschungen dürften die aus morphologischen und geologischen Gründen abgeleitete Annahme, daß es sich bei den Hohlformen um Erdfälle handele, erheblich stützen.
Literatur und Archivquellen
ANDERSEN, S. Th. (1973): The differential pollen productivity of trees and its significance for the Interpretation of a pollen diagram from forested region. - In: BIRKS, H. J. B. & WEST, R. G.: Quaternary plant ecology 1973: 109-115. Berlin: Blackwell.
AVERDIECK, F.-R. (1971): Zur postglazialen Geschichte der Eibe (Taxus baccata L.) in Nordwestdeutschland. - Flora 160: 28-42. Jena.
AVERDIECK, F.-R. (1986): Probleme der Palynologie an einem Erdfall. - Documenta naturae 33: 8-10. München.
BECKHAUS, K. (1893): Flora von Westfalen. - Münster: Aschendorff. (1096 S.)
BUDDE, H. (1951): Die Trocken- und Halbtrockenrasen und verwandte Gesellschaften im Wesergebiet bei Höxter (eine pflanzengeographische Untersuchung). - Abh. Landesmus. Naturk. Münster 14 (3). Münster (Westf). (38 S.)
BURRICHTER, E. (1970): Beziehungen zwischen Vegetations- und Siedlungsgeschichte im nordwestlichen Münsterland. - Vegetatio 20: 199-202. The Hague.
BURRICHTER, E. (1976): Vegetationsräumliche und siedlungsgeschichtliche Beziehungen in der Westfälischen Bucht. Ein Beitrag zur Entwicklungsgeschichte der Kulturlandschaft. - Abh. Landesmus. Naturk. Münster 38 (1): 3-14. Münster.
BURRICHTER, E. & POTT, R. (1983): Verbreitung und Geschichte der Schneitelwirtschaft mit ihren Zeugnissen in Nordwestdeutschland. - Tuexenia 3 (Festschrift Ellenberg): 443-453. Göttingen.
ERDTMAN, G. (1954): An introduction to pollen analysis. - Mass. (U. S. A): Waldham. (239 S.)
FIRBAS, F. (1949): Spät- und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen. Bd. 1: Allgemeine Waldgeschichte. - Jena. (480 S.)
FIRBAS, F. (1952): Spät- und nacheiszeitliche Waldgeschichte Mitteleuropas nördlich der Alpen. Bd. 2: Waldgeschichte der einzelnen Landschaften. - Jena. (256 S.)
GIGAS, J. (1672): 3 Karten "Dioecesis Paderbornensis". In: FERDINAND VON FÜRSTENBERG (1713): Monumenta Paderbornensis. 3. Aufl. - Frankfurt und Leipzig. (309 S.)
GRUPE, O. (1929a): Geologische Karte von Preußen l : 25000. 2. Aufl. Blatt Höxter. Erläuterungen. - Berlin. (36 S.)
GRUPE, O. (1929b): Geologische Karte von Preußen l : 25000. 2. Aufl. Blatt Holzminden. Erläuterungen. - Berlin. (71 S.)
GUTHEIL, H. E. (1837): Beschreibung der Wesergegend um Höxter und Holzminden. Holzminden. (76 S.)
HAEUPLER, H. (1976): Atlas zur Flora von Niedersachsen. - Scripta geobot. 10. Göttingen. (376 S.)
HAVINGA, A. (1962): Een palynologisch onder-zoek van in dekzant ontwikkelde bodemprofielen. - Proefschrift Wageningen. (165 S.)
HESMER, H. (1928): Die Waldgeschichte der Nacheiszeit des nordwestdeutschen Berglandes auf Grund von pollenanalytischen Mooruntersuchungen. - Z. Forst- u. Jagdwesen 1928: 1-67. Berlin.
JÜNGST, L. V. (1852): Flora Westfalens. -Münster. (438 S.)
KLEINAU, H., PITZ, E., VORTHMANN, A. & REINWALD, E. (1977): Der Solling im 3. Viertel des 18. Jahrhunderts. Karte l : 25000. - Veröff. Histor. Kommission Nieder. S. 23. Wolfenbüttel.
KNAPP, G., (1983): Geologische Karte von Nordrhein-Westfalen 1 : 25000, Blatt 4321 Borgholz. Erläuterungen. - Krefeld. (160 S.)
KNÖRZER, K. H. (1949): Die Vegetation des Torfmoores im Solling und die nacheiszeitliche Waldgeschichte dieses Gebirges auf Grund der Pollenuntersuchungen. - Staatsexamensarbeit Göttingen. (Unveröff). (Vgl. FIRBAS, F. 1952: 75-84).
KRABBE, G. (1603): Solling-Karte ca. 1 : 22000. - Nieders. Staatsarchiv K 0203. Wolfenbüttel.
LEESCH, W. (1961): Inventar des Archivs der Stadt Höxter, XII (II Dokumente). - Archive Westfalens NF 1. Münster. (592 S.)
LESEMANN, B. (1968): Pollenanalytische Untersuchung eines Flachmoores im Umlauftal der Weser bei Bodenfelde. - Ber. Naturhist. Ges. Hannover 112: 91-96. Hannover.
MERKEL, E. (1930): Die Geschichte des Corveyer Waldes. - Neuherausg. u. red. von PREYWISCH, K. (1978) Egge-Weser, Sonderveröff. 1. Höxter. (241 S.)
MERKT, J. & STREIF, H. (1970): Stechrohr-Bohrgeräte für limnische und marine Lockersedimente. - Geol. Jb. 88: 137-148. Hannover.
NIETSCH, H. (1955): Untersuchungen über die jüngere Talgeschichte der Weser bei Schlüsselburg und das Alter des Niederterrassenlehms bei Stolzenau. - Jb. geograph. Ges. Hannover 1954 u. 1955: 19-28. Hannover.
OBERDORFER, E. (1979): Pflanzensoziologische Exkursionsflora. 4. Aufl. - Stuttgart: Ulmer. (997 S.)
OVERBECK, F. TH. (1975): Botanisch-geologische Moorkunde unter besonderer Berücksichtigung der Moore Nordwestdeutschlands als Quellen zur Vegetations-, Klima- und Siedlungsgeschichte. - Neumünster. (719 S.)
POTT, R. (1982): Das Naturschutzgebiet "Hiddeser Bent - Donoper Teich" in vegetationsgeschichtlicher und pflanzensoziologischer Sicht. - Abh. Westf. Mus. Naturk. Münster 44 (3) Münster. (108 S.)
POTT, R. (1984): Pollenanalytische Untersuchungen zur Vegetations- und Siedlungsgeschichte im Gebiet der Borkenberge bei Haltern in Westfalen. - Abh. Westf. Mus. Naturk. Münster 46 (2). Münster. (28 S.)
POTT, R. (1985a): Vegetationsgeschichtliche und pflanzensoziologische Untersuchungen zur Niederwaldwirtschaft in Westfalen. - Abh. Westf. Mus. Naturk. Münster 47 (4). Münster. (75 S.)
POTT, R. (1985b): Beiträge zur Wald- und Siedlungsentwicklung des Westfälischen Berg- und Hügellandes auf Grund neuer pollenanalytischer Untersuchungen. - Siedlung u. Landschaft 17: 1-38. Münster.
POTT, R. (1990): Historische Waldnutzungsformen Nordwestdeutschlands. - Heimatpflege in Westfalen 3 (2): 1-9. Münster.
PREYWISCH, K. (1962): Zur Waldgeschichte des Naturschutzgebiets "Ziegenberg". - Höxter-Corvey 10 (3): 1-2. Höxter.
RAUS, T. (1977): Exkursionen der Floristisch-soziologischen Arbeitsgemeinschaft während der Jahrestagung in Höxter 1976. - 50 Jahre Floristisch-soziologische Arbeitsgemeinschaft (1927-1977). - Mitt. flor.-soz. Arbeitsgem. N.F. 19/20: 437-448. Todenmann, Göttingen.
RODE, L. (1844): Allgemeine Beschreibung des Communalforstes der Stadt Höxter. - Stadtarchiv Höxter B XV. (27 S.)
ROHDE, U. (1983): Die Pflanzengesellschaften des Naturschutzgebietes "Ziegenberg" bei Höxter. - Diplomarb. Univ. - GH Paderborn, Abt. Höxter. (Unveröff.)
RUNGE, F. (1972): Die Flora Westfalens. 2. Aufl. - Münster: Westf. Vereinsdruckerei. (550 S.)
RUNGE, F. (1973): Die Pflanzengesellschaften Deutschlands. 475. Aufl. - Münster: Aschendorff. (246 S.)
SCHNEEKLOTH, H: (1967): Vergleichende pollenanalytische und 14C-Datierungen an einigen Mooren im Solling. - Geol. Jb. 84: 717-734. Hannover.
SCHÜTRUMPF, R. (1981): Der pollenanalytische Nachweis einer schwimmenden Insel" und die Anzahl der Siedlungsphasen am mittelsteinzeitlichen Fundplatz Duvensee, Kr. Herzogtum Lauenburg/Schleswig-Holstein. Kölner Jb. 15 (1975-1977), Festschrift Schwabedissen: 161-180. Berlin.
SIES (SIEHS), F. H. (1704): "Die Schlacht an der Brunsburg". Gemälde, Öl auf Leinwand, ca. 155x210 cm. Rathaus Höxter.
STECKHAN, H.-U. (1961): Pollenanalytisch-vegetationsgeschichtliche Untersuchungen zur frühen Siedlungsgeschichte im Vogelsberg, Knüll und Solling. - Flora 150: 514-551. Jena.
STEPHAN, H.-G. (1978): Archäologische Studien zur Wüstungsforschung im südlichen Weserbergland I (Text). - Hildesheim. (293 S.)
WILLERDING, U. (1960): Beiträge zur jüngeren Geschichte der Flora und Vegetation der Flußauen (Untersuchungen aus dem Leinetal bei Göttingen). - Flora 149: 435-476. Jena.
WILLERDING, U. (1965): Urgeschichtliche Siedlungsreste in Rosdorf, Krs. Göttingen. II. Die Pflanzenreste der Bandkeramischen Siedlung. - Neue Ausgrabungen u. Forschungen in Nieders. 2: 44-60. Hildesheim.
WILLERDING, U. (1968): Beiträge zur Geschichte der Eibe (Taxus baccata L.). - Presse-Archiv 3: 96-155. Göttingen.
WILLERDING, U. (1971): Ergebnisse vegetationsgeschichtlicher und paläo-ethnobotanischer Untersuchungen im südlichen Niedersachsen. - Göttinger Jb. 19:5-20. Göttingen.
WILLKOMM, H. (1995): Radiokohlenstoffdatierung der Sedimente der Grundlosen . - Veröffentlichungen des Naturkundlichen Vereins Egge - Weser 7: 79-85. Höxter
Anschrift der Verfasser:
Dr. Fritz-Rudolf Averdieck, Institut für Ur- und Frühgeschichte, Neue Universität, Bau 17, 24098 Kiel Kurt Preywisch, Ansgarweg 19, 37671 Höxter
Tabelle 1: Pollenfunde, die in dem Diagramm in Summenkurven dargestellt sind. Die Buchstaben hinter den Pflanzennamen bedeuten den stratigraphischen Abschnitt, z. B. a = 1%-10% im Abschnitt a enthalten, a! = mehr als 10%, (a) = weniger als 1%. Eine Zahl hinter dem Buchstaben bedeutet den Maximalwert, bezogen auf die Gesamt-Baumpollensumme.
1. Sonstige Kulturanzeiger
Chenopodiaceae k 4,5; i; (h); (f); (d); c; (b) Polygonum aviculare k 3,2; i; (h); (c); (b) Polygonum convolvulus (k); (e); (b) Scleranthus annuus k 1,6; i; (g) Spergula k 2,6; i; (f); c; (b) Spergularia i 2,3; (c) Trifolium arvense-Typ (i); (c); (b) Trifolium pratense k 1,6; i; (h); (e); (c); (b)
2. Sonstige Pflanzen vorwiegend trockener Standorte
Anemone-Typ (k); i 1,3; h 1,3; (g); (e); (d); (c); b cf. Campanula (k); (g); (c) Coronilla-Typ (h); (e) Echium-Typ (k) Linum catharticum i 1,1 Lycopodium clavatum (k) Mercurialis annua (d) Mercurialis perennis (k); (f); (c) Polypodium vulgäre i 1,1; (h); (g); (f); (d); (c); (b) Pteridium aquilinum k; i 5,9; h; (g); f; (e); d; c; b; (a) Scleranthus perennis (i) Silene-Typ i 1,0; (f); (c) Stellaria holostea (k) Teucrium cf. scorodonium (g); (e)?; (c)
3. Sonstige Pflanzen indifferenter Standorte
cf. Allium c 1,5; (b) Balsaminaceae (g); (e); c 3,6 Boraginaceae indet. i 1,1 Capsella/Sinapis-Typ k! 25,0; h; g; f; (e); c; b; (a) Cardamine-Typ k 5,0; i; h; f; (e); (d); c; (b) Cerastium-Typ (h); (c); (b) Epilobium i 1,0 Eupatorium-Typ k! 11,3; i; h; (g); (f); c Chaerophyllum temulum h 2,3; f; d; (c) Cicuta-Typ k!; i; h; g!; f; e!; d! 29,3 Ficaria-Typ i 1,9; (h); (g); (e); (d); (c); (b) cf. Galium k 4,1; i; h; g; f; e; d; c; b; Geranium (g) Geum-Typ (k); i 2,0; (e); (d); (c) Hypericum (i); (g); (f); e 2,1; d; c; (b) Labiatae, 3-colpat (h); (g); e; (f); (d); c 1,5 Labiatae, 6-colpat (k); i 1,0; (h); (g); (f); (e); (c) Lamium-Typ k 1,3; h Lysimachia vulgaris-Typ (g); (c); (b) Melampyrum i 1,0; (h); (b) Melandrium rubrum (g); (f); (e)
Melilotus-Typ k 1,1; (g); (f); (c); (b)
cf. Moehringia (c)
Oenanthe-Typ k!; i! 62,9; h!; g; f!; e!; d; c!; b
cf. Pedicularis (b)
Polygonom hydropiper-Typ i 1,0
Potentilla Comarum-Typ i; (h); g; f 1,7; e; (d); c; (b)
Ranunculus cf. acer (f); (b)
Ranunculus bulbosus/
repens-Typ (i); (g); (f); (c)
Ranunculus indet. (k); i 1,6; (g); (f); (c); (b)
cf. Rhinanthus (i); (g); (f); (e); c; b 1,1; (a)
Rosaceae indet. (f); (d)
cf. Scabiosa c 1,0
cf. Scrophularia nodosa k 2,2; i; (h); (g)
cf. Scutellaria (h); (g); (d); (c)
Solanum cf. nigrum (d); (c)
Succisa i 1,0; (g); (f); (b)
Thalictrum (c); (b)
Trifolium indet. (i); (f); (e); c 1,6
Trifolium cf. repens k 1,7; i; h; f; (e); (d); c; (b)
Umbelliferae indet.,
hantelförmiger Typ k!; i! 50,5; h!; g; f!; e!; c; b; a
Umbelliferae indet.,
ovaler Typ k 6,3; i; h; (g); f; e; (d); c; b; (a)
Veronica-Typ (k); (i); (h); (g); (f); (e); (c); (b); (a)
4. Sonstige Pflanzen vorwiegend feuchter Standorte
Alisma k; i; (h); g; f; e; d; c 4,9; (b) Caltha k 1,3; i Dryopteris filix-femina (g) Filipendula k 7,5; i; h; g; f; e; d; c; b; (a) Hippuris (e) Humulus/Cannabis-Typ (k); (i); h 6,8; (g); (c) Lotus uliginosus (k); (h) Lycopodium annotinum (i) Lysimachia thyrsiflora (g); (d); (c) Lythrum (h); e 2,7; (d); (c) Menyanthes (d); (b) Ophioglossum (k) Parnassia (g); (c) Ranunculus cf. lingua k 3,8; (i); (h); (f) Rumex cf. hydrolapathum (k); i 4,4; (g) Solanum dulcamara k; i!; h! 22,1; g; (f); (e) Thelypteris palustris (h); (g); (f); d 7,6; c Urtica k; i! 29,7; h; g; f; e; d; c!; (b); (a) Valeriana cf. officinalis (g); (c) cf. Viola palustris (k); (e); (c); (b)
5. Schwimmblatt- u. submerse Pflanzen
Batrachium div. spec. k 8,8; i; h; (g); f; e; (d); c; b Callitriche k; i; h; g; f; e 9,0 Hottonia k!; i; h; g; f! 11,1; e; (d); c; (b) cf. Hydrocharis (k); (f) Lemna k; i; h; g!; f!; e!; d; c! 66,8; (b) Nymphaea (c) Potamogeton pectinatus-Typ (d); (c) Potamogeton/Triglochin-Typ k!; i; (h); (g); f!; e; d! 46,4; c; b; a
Tabelle 2: Früchte- u. Samenfunde, die nicht im Diagramm aufgeführt worden sind. Es handelt sich dabei um vereinzelte Vorkommen, selten mehr als eines je Abschnitt. Unsichere Bestimmungen sind in Klammern gesetzt.
Acer cf. campestre g 2; g l Brassica/Sinapis g 3;g l;c cf. Cerastium (k); (i); (c) Compositae indet. k Crataegus c Cruciferae indet. i Epilobium cf. hirsutum i; c Eupatorium cannabinum k Fagus (g 1) Filipendula d Fraxinus excelsior g 3 Galium cf. palustre c Hypericum humifusum k Iris pseudacorus (h); g 3; (f); (e), d Lapsana g 3 Mentha cf. aquatica i; c Oxalis acetosella d Polygonum cf. persicaria i; h; c Prunella vulgaris c Quercus (1) Rosaceae indet. c Sambucus nigra g 2; f; c Sambucus racemosa h; c Scirpus lacustris (g 2) Scleranthus c Umbelliferae indet. g 3; b
Tabelle 3: Gegenüberstellung der Siedlungsfluktuationen nach den Pollendiagrammen aus den Grundlosen und den benachbarten östlichen Bergländern Westfalens und des Sollings, vorzugsweise angelehnt an die 14C-Daten und an die mit ihnen verknüpften Leithorizonte von STECKHAN 1961, SCHNEEKLOTH 1967
|
Östliches Westfälisches Bergland (Teutoburger Wald, Eggegebirge) (POTT 1985) |
Grundlose bei Höxter (AVERDIECK, PREYWISCH & WILLKOMM) |
Solling (SCHNEEKLOTH 1967, unter Einbeziehung von KNÖRZER 1949 und STECKHAN 1961) |
| 1. Hälfte 15. Jahrhundert starke Siedlungsausweitung | ||
| Um 1300 Wüstungsperiode | Spätestens ab 1100 starke Entwaldung | 11/12. Jahrhundert Beginn großer Rodungen |
| Vor 1100 kurze Siedlungsdepression | ||
| 600 - 1300 sächsische und fränkische Landnahme, ab 800 starke Rodungen | Zwischen 600 u. 1100 (9. Jahrh. ?) Siedlungsexpansion | Nach 700 Getreideanbau |
| Zwischen 300 u. 600 (eher 600 ?) verstärkte Siedlungstätigkeit | ||
| 100 - 450 Siedlungsdepression, Völkerwanderungszeit | Nach 60 oder 150 - 700 Siedlungsleere | |
| 100 v. Chr. - 300 oder eher 600 n. Chr. starke Siedlungsdepression | ||
| 0 - 100 Siedlungsbelebung Um Chr. Geb. Siedlungsdepression |
Vor 60 - nach 150 erhöhte Siedlungsanzeichen | |
| 850 v. Chr. - vor 60 oder 150 n.Chr. Siedlungsrezession | ||
| 1300 v. Chr. - um Chr. Geb. spätbronze-früheisenzeitliche Besiedlung | Vor 1000 v. Chr. - 100 v. Chr. (vermutlich) kontinuierliche bronze-früheisenzeitliche Besiedlung | 1250 v. Chr. - 850 v. Chr. erhöhte Kulturanzeichen, aber kaum Getreide |
| 1300 v. Chr. - 1200 v. Chr. Depression der Mittleren Bronzezeit | ||
| 2000 v. Chr. - 1250 v. Chr. Siedlungsrezession | ||
| 2400 v. Chr. - 2000 v. Chr. erste Kulturanzeichen | ||
| 4000 v. Chr. - 1200 v. Chr. frühbronzezeitliche Besiedlung |
Tabelle 4: Pollenprozente aus den Rezentproben. Berechnungsweise wie im Pollendiagramm. Bei Probe 5 sind Stückzahlen (in Klammern) aufgeführt. In Rubrik "Sonstige Kräuter trockener Standorte" setzt sich in Probe 1 die Zahl aus vielen Arten zusammen, in Probe 2 ist Ranunculus cf. repens mit 20 % beteiligt. Die Werte der Sumpf- u. Wasserpflanzen werden in den Proben 3 und 5 fast ausschließlich von Callitriche gebildet. Pollenerhaltung: + gut 0 mäßig - schlecht
