Die Schweineweide als Lebensraum für Vögel (Aves)
und Laufkäfer (Coleoptera: Carabidae),
dargestellt am Beispiel des Echeler Bruches in der Warburger Börde (Kreis Höxter)
Von Benjamin T. Hill, Burkhard Beinlich, Heiko Köstermeyer und Lothar Beck
Zusammenfassung
Das Verbundforschungsvorhaben “Schweinefreilandhaltung im Rahmender Landschaftspflege“ hat sich in den Jahren 1999-2003 mit der Frage beschäftigt, inwieweit die extensive Haltung von Hausschweinen einen Beitrag zum Erhalt gefährdeter Offenlandstandorte leisten kann, bzw. ob und in welchem Umfang gefährdete Arten von der Wühltätigkeit der Tiereprofitieren. Am Beispiel einer bis zur Einrichtung der Schweineweide brachliegenden Niedermoorfläche in der Warburger Börde(Kr. Höxter, Ostwestfalen) werden die Auswirkungen der Schweinebeweidung auf tierische Lebensgemeinschaften am Beispiel der Vögel und Laufkäfer erörtert und die Wertigkeiten der Zönosen im Vergleich zu benachbarten Brachflächen bzw.konventionell genutzten Feuchtgrünländern diskutiert.
Trotz gewisser Einschränkungen aufgrund der relativ geringen Flächengrößen der Untersuchungsfläche und der damit einhergehenden Randeffekte sowie fehlender Versuchswiederholungen könnenbezüglich der Avifauna folgende Schlussfolgerungen gezogen werden: Extensiv genutzte Schweineweiden in Feuchtgebieten bieten ganzjährig geeignete Habitate und Strukturen für Vogelarten des Offenlandes, der Feuchtwiesen und der Röhrichte bzw. Hochstaudenfluren. Aufgrund der engen Verzahnung unterschiedlichster Strukturen geschieht dies selbst auf vergleichsweise kleinen Flächen in einem Umfang, wie es von anderen landwirtschaftlichen Nutzungen nicht gewährleistet wird. Insbesondere zur Förderung der in Mitteleuropa stark gefährdeten Wiesenbrüter stellt die Schweinebeweidung in Abhängigkeit von den standörtlichen Parametern eine geeignete Nutzungsform dar.
Bezüglich der Laufkäferfauna ist festzuhalten, dass die Artengemeinschaft in erster Linie von der Bodenfeuchte bzw. der Dauer der Überstauung bestimmt wird – dies spiegelt sich auch bei der Betrachtung ausgewählter funktioneller Merkmale wider. Erst an zweiter Stelle wirkt sich die Nutzung der Flächen auf die Zusammensetzung der Lebensgemeinschaft aus. Insbesondere die Strukturparameter "Offenbodenanteil" und "Vegetationshöhe/-dichte" beeinflussen die Laufkäferzönosen maßgeblich.
Die Schweineweide wies mit 41 nachgewiesenen Laufkäferarten im Vergleich zu benachbarten Brachen (36 Arten) eine höhere Diversität auf– die Käfer zeigten dort ebenfalls höhere Aktivitätsdichten. Typische Feuchtgrünlands- bzw.Niedermoorarten waren überwiegend auf den lange überstauten,nassen Teilbereichen der Untersuchungsflächen anzutreffen,wobei mehrere Arten – darunter der seltene Elaphrusuliginosus – Präferenzen für die Schweineweide zeigten. Demgegenüber drangen auf die frisch-feuchten Teilbereiche vermehrt euryöke Grünlands- und Ackerarten vor.
Der Vergleich mit nahe gelegenen, als Wiesen genutzten Niedermoorresten ergab nur sehr geringe Übereinstimmungen. Hierfür dürfte die unterschiedliche Nutzungsgeschichte, aber auch die verschiedene Erfassungsmethodik eine Rolle spielen.
Insgesamt kann aus Sicht der untersuchten Tiergruppen die neu eingeführte extensive, saisonale Schweinebeweidung im Bereich des degradierten Niedermoores als erfolgreich bewertet werden. Die charakteristischen Artengemeinschaften konnten erhalten bzw. neu etabliert werden, darüber hinaus profitierten einige selten bzw. gefährdete Arten von der neuen Nutzungsform.
98
Wichtige Ziele des Naturschutzes, wie die Förderung der Biodiversität oder die Schaffung geeigneter Lebensräume für gefährdete Arten, konnten somit realisiert werden.
1 Einleitung
Niedermoore gehören in Deutschland zu den Biotoptypen, die im vergangenen Jahrhundert erhebliche Bestandseinbrüche zu verzeichnen hatten (Riecken et al. 1994). So haben im norddeutschen Tiefland Meliorationsmaßnahmen zu einer Überführung der Feucht- und Nasswiesen in produktivere mesophile Grünländer geführt (IRMLER et al. 1998). Demgegenüber ist in den Mittelgebirgsregionen – wo landwirtschaftlich genutzte Niedermoore eher kleinflächig in Bach- und Flusstälern, Muldenlagen und an Sonderstandorten auftraten – in erster Linie die Nutzungsaufgabe als Gefährdungsfaktor wirksam. Im Verlaufe der Sukzession werden die ehemals wertbestimmenden Sumpfdotterblumenwiesen, Kleinseggensümpfe und Großseggenrieder durch artenarme Hochstaudengesellschaften verdrängt oder entwickeln sich zu Landröhrichten, die nur einen Bruchteil der ehemaligen naturschutzfachlichen Wertigkeit aufweisen.
Von Seite des Naturschutzes wurde deshalb lange Zeit versucht, über Pflegemaßnahmen diese Entwicklung zu verhindern bzw. zu verzögern. Aufgrund der meist hohen Grundwasserstände ist eine maschinelle Pflege aber mit großen Schwierigkeiten verbunden. Zusammen mit der allseits schlechten Finanzsituation der öffentlichen Hand und einer z. T. fehlenden Zielbestimmung seitens des Naturschutzes hat dies vielerorts zu einem deutlichen Pflegerückstand in den betroffenen Lebensräumen geführt. Der Entwicklung und Umsetzung neuartiger/alternativer Methoden zum Erhalt solch marginaler Grünlandstandorte kommt deshalb eine hohe aktuelle Bedeutung zu. Verstärkt wird dabei auf extensive Weidesysteme gesetzt (z.B. CONRADI & KRÜGER 1999; KAMPF 2000; RIECKEN et al. 2001). Ein bisher völlig neuer Aspekt ist der mögliche Einsatz von Hausschweinen zum Erhalt des Feucht- und Nassgrünlandes (BEINLICH 1998; HILL et al. 2000). Erste Ergebnisse legen nahe, dass eine extensive, saisonal betriebene Schweinebeweidung sowohl auf trockenen Grünlandbrachen als auch in Bach- und Flussauen positive Auswirkungen auf die Laufkäfer entfaltet (HEROLD 2001; HILL et al. 2003). Für Niedermoorstandorte sind bislang keine zooökologischen Untersuchungen erfolgt – die Ergebnisse von NEUGEBAUER (2003) für die Vegetation zeigen aber positive Entwicklungen auf.
Der vorliegende Artikel will am Beispiel eines kleinen degradierten Niedermoorrestes inmitten einer intensiv landwirtschaftlich genutzten Beckenlandschaft (Warburger Börde) aufzeigen, inwieweit die neue Nutzungsform die Artengemeinschaften ausgewählter Tiergruppen (Vögel, Laufkäfer) beeinflusst. Zur Eichung der Ergebnisse dienen Brachflächen im Echeler Bruch sowie im benachbarten Rietbruch. Nach Vorgaben des Vertragsnaturschutzes bewirtschaftete Wiesen, ebenfalls im Rietbruch gelegen, dienen als Referenz. Weiterhin werden Literaturangaben aus einem weiteren Niedermoor in räumlicher Nähe zu den beiden anderen Standorten (NSG „Körbecker Bruch“) berücksichtigt.
Folgende Fragestellungen stehen im Mittelpunkt dieses Artikels:
Wie reagieren die Brutvögel auf die Schweinebeweidung?
Wird das Gebiet für rastende Limikolen attraktiver?
Wie entwickelt sich die Laufkäfergemeinschaft der Schweineweide im Unterschied zu den brach gefallenen Flächen? Können lebensraumtypische und ggf. seltene/gefährdete Arten erhalten/gefördert werden?
Wie verändern sich für die Laufkäferbesiedlung bedeutsame strukturelle Qualitäten der Flächen?
Ergeben sich Unterschiede bei den funktionellen Merkmalen der Laufkäfer?
Wie sind die Laufkäferzönosen in nahe gelegenen Niedermoorresten strukturiert? Ist die herkömmliche Nutzung (in diesem Falle Mahd) besser geeignet, die typischen Arten zu erhalten?
99
2 Untersuchungsgebiete und Methodik
Die Untersuchungsflächen liegen in der Warburger Börde südwestlich von Borgentreich, Kreis Höxter, NRW (Abb. 1, 2). Sie befinden sich alle in einer Höhe von 170 bis 210 m ü. NN. Während der Echeler Bruch nur eine Größe von ca. 10 ha aufweist, sind Rietbruch ca. 40 ha und Körbecker Bruch knapp 100 ha groß. Bei den Böden handelt es sich an allen drei Standorten um Niedermoor bis Erdniedermoor bzw. Moorgley bis Anmoorgley, jeweils aus Niedermoortorfen hervorgegangen (JAHN 2003, schriftliche Mitteilung).
| Abb. 1: | Lage der Untersuchungsflächen Echeler Bruch (EB), Rietbruch (RB) und Körbecker Bruch (KB) (Karte: W. KÖBLE; © Geobasisdaten: LVA NRW, Bonn, 2005) |
a. Echeler Bruch (EB)
Es handelt sich um eine ehemals drainierte und kurzzeitig auch als Acker genutzte, ca. 10 ha große Niedermoorfläche, die seit Ende der 1980er Jahre brach liegt. Zwei größere Gräben durchziehen das Gebiet mittig, weitere Gräben entwässern das Niedermoor randlich (Abb. 2). Aufgrund fehlender Grabenpflege sind die zentralen Gräben weitgehend verlandet, sodass der Grundwasserstand im zentralen Bereich zwischenzeitlich wieder knapp unter Flur liegt, im Winterhalbjahr sogar wenige cm darüber. Die Fläche der 2 ha großen Schweineweide fällt nach Nordosten, in Richtung der Gräben ab, sodass sich standörtlich hinsichtlich der Feuchtigkeit klare Unterschiede ergeben – die Bereiche zwischen beiden Gräben sind bis weit in das Frühjahr hinein flach überstaut. Im Folgenden wird dieser Bereich als „nass“ und die höher gelegenen als „frisch-feucht“ bezeichnet.
100
Die Beweidung der überwiegend dichten Hochstaudenfluren und eingestreuten Seggen- und Binsenbestände wurde im Winterhalbjahr 1999/2000 aufgenommen. Eine standörtlich vergleichbare, direkt angrenzende Brachfläche wurde in die Laufkäferuntersuchungen als Referenz einbezogen (vgl. Abb. 2). Für die Untersuchungen der Laufkäfer ergeben sich demnach 4 Teilflächen mit unterschiedlicher Nutzung (Schwein und Brache) bzw. Bodenfeuchte (frisch-feucht und nass).
Die avifaunistischen Untersuchungen wurden auf der gesamten Fläche durchgeführt.
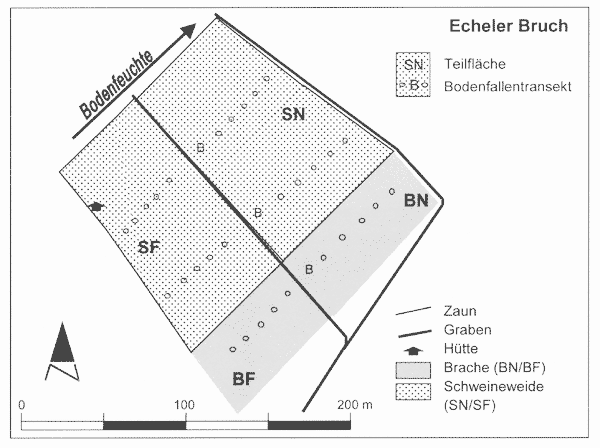
| Abb. 2: | Detailplan der zentralen Untersuchungsfläche Echeler Bruch mit Lage der vier Teilflächen, die sich nach Bodenfeuchte (Nass + Frisch-Feucht, s. Angabe des Bodenfeuchte-Gradienten) und Nutzung (Schweineweide + Brache) differenzieren. (Karte: Autoren) |
Die Beweidung erfolgte, abgesehen vom ersten Jahr, jeweils im Spätsommer ab August für 8-12 Wochen. Es wurden ca. 15-20 Läuferschweine / ha (< 0,3 GV/ha) aufgetrieben. Bei der eingesetzten Rasse handelte es sich um Düppeler Weideschweine (s. Abb. 9, 10).
Nach dreijähriger Beweidung wird die Vegetation aus einem Mosaik von hochwüchsigen Dominanzbeständen aus Wiesenfuchsschwanz (Alopecurus pratensis), Glatthafer (Arrhenaterum elatius) oder Weidenröschen-Hochstaudenfluren (Epilobium spp.) sowie Urtica dioica- und Calamagrostis epigejos-Beständen im Wechsel mit niedrigwüchsigeren Carex disticha-, Caltha palustris- und Ranunculus flammula/sceleratus-Beständen gebildet (vgl. Neugebauer 2003). Nach Abtrieb der Weideschweine im Herbst/Winter finden sich über mehrere Monate größere Rohbodenbereiche (bis zu 30 % Flächenanteil) auf den Weiden.
b. Rietbruch (RB)
1 km südlich des Echeler Bruchs befindet sich die etwa 40 ha große Niedermoorfläche des Rietbruches, die sich südöstlich im Rösebecker Bruch fortsetzt. Auch diese Fläche ist meliorisiert und wird überwiegend als gemähtes Grünland genutzt. Die zentralen, feuchteren Bereiche liegen brach und weisen Landröhrichte auf. Im Un-
101
c. Körbecker Bruch (KB)
Das NSG liegt östlich von Echeler Bruch und Rietbruch in gut 2 km Entfernung. Die trockeneren Bereiche werden durchgehend als Mähwiesen ohne Düngung extensiv genutzt (1. Mahd jeweils nach dem 15.06.), die zentralen Feuchtbereiche liegen zum Teil brach und weisen Hochstaudenfluren bzw. Röhrichte auf (LOHR 2005, in diesem Heft). Gehölze beschränken sich auf die randlichen Bereiche des Schutzgebietes, welches im Kreis Höxter das bedeutendste Wiesenbrütergebiet darstellt.
Die Erfassung der Brutvögel erfolgte in den Jahren 2000 bis 2002 durch flächendeckende Begehungen im Zeitraum von März bis Juli (insgesamt 7, zur Methodik der Revierkartierung vgl. BIBBY et al. 1995). Der Nachweis des Wachtelkönigs erfolgte im Rahmen von abendlichen und nächtlichen Begehungen. Zusätzlich zu den Brutvögeln wurden weiterhin zu den Zugzeiten (Frühjahr, Herbst) die Rastvögel kartiert – es fanden jeweils zwischen 6 bis 8 Begehungen/Jahr statt. Für den Echeler Bruch stehen außerdem eigene Daten, die mit gleicher Methodik erhoben wurden, aus den Jahren 1998 und 1999 zur Verfügung.
Im Rietbruch wurde die Avifauna nach gleicher Methodik im Jahr 2002 erhoben. Für dieses Gebiet liegen weiterhin Daten aus den Jahren 1997 von SCHLEEF & MI&SZLIG;FELDT (1998) sowie eigene, nach vergleichbarer Methodik erhobene Daten aus den Jahren 2000 und 2001 vor.
Avifaunistische Daten zum Körbecker Bruch wurden, ebenfalls nach gleicher Methodik, im Jahr 2002 erhoben. Für dieses Gebiet sind weiterhin Daten aus dem Jahr 1998 (FIGURA et al. 1999) und 1981 (SMOLIS 1982) verfügbar.
Die Erfassung der Laufkäfer erfolgte mit Bodenfallen in Anlehnung an BARBER (1931). Aufgrund des ausgeprägten Erkundungstriebs der Schweine in Kombination mit dem starken Drang zum Wühlen machte die Entwicklung eines speziellen Fallentyps notwendig: die Fanggläser wurden in 30 cm lange PVC-Rohre (Durchmesser 12 cm) eingelassen (vgl. MANDERBACH 1998, S. 35) und zu einem Drittel mit gesättigter Kochsalzlösung (mit einigen Tropfen Entspannungsmittel) gefüllt. Insgesamt wurden im Echeler Bruch auf der Schweineweide zwei, auf der Brache eine Reihe(n) mit je 10 Fallen senkrecht zu den Gräben aufgestellt, um die standörtliche Heterogenität abzubilden (Abstand ca. 10 m). Für den direkten Vergleich der unterschiedlichen Nutzung (Brache – Schweineweide) wurde aber nur eine dieser Reihen berücksichtigt (s. Kap. 3.2.2). Die Fangzeiträume wurden zum einen stark von der Hydrologie des Gebietes (z. T. hohe Grundwasserstände im Frühjahr), zum anderen vom Brutgeschäft des Wachtelkönigs (eingeschränkte Betretungserlaubnis) bestimmt. Deshalb erfolgte nur eine extensive Beprobung mit 3 Fangdurchgängen im Frühsommer, Sommer und Herbst (jeweils 10-14 Tage). Zudem stellte sich heraus, dass die getroffenen Vorsichtsmaßnahmen zum Abhalten der Schweine nicht ausreichend waren – es kam immer wieder zu Fallenverlusten.
Im Echeler Bruch wurde 2000 an insgesamt 4321, 2001 an 379 und 2002 an 370 Fallentagen gefangen. Bedingt durch die Projektlaufzeit (Beginn im Herbst 1999) war keine Nullaufnahme möglich. Im RB wurden im Jahr 2001 insgesamt 10 Fallen zu den gleichen Zeiten aufgestellt (252 Fallentage).
Die Bestimmung der Käfer erfolgte nach (FREUDE et al. 1976; TRAUTNER & GEIGENMÜLLER 1987; LOMPE 1989). Eine Unterscheidung der Agonum afrum/viduum-Gruppe sowie von Trechus quadristriatus/obtusus erfolgte nicht. Die Nomenklatur richtet sich nach TRAUTNER et al. (1997).
________________________
1Insgesamt betrug die Fangdauer 780 Tage, in die vergleichende Analyse wurden aber nicht alle Fallen mit einbezogen (s. Text).102
3 Ergebnisse
3.1 Vögel
3.1.1 Brutvögel
Im gesamten Echeler Bruch wurden zwischen 1998 und 2002 zwischen 17 und 21 Brutvogelarten nachgewiesen. Diese Arten umfassen die unterschiedlichsten Gilden, von Hecken- und Waldbewohnern über Vögel der Feldflur bis hin zu Wasservögeln. Da es sich bei der Schweineweide um Grünland mit eingestreuten Röhrichten und Hochstauden handelt, werden im Folgenden nur die zwei Gilden der Wiesenbrüter und die der Arten der Röhrichte und feuchten Hochstaudenfluren weiter betrachtet.
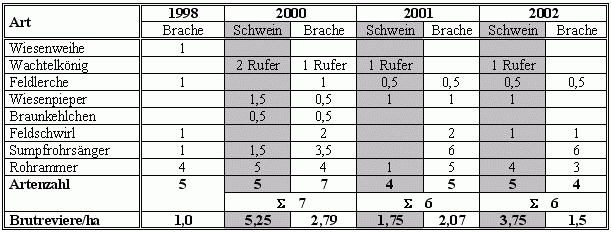
In Tab. 1 ist der Ausgangsbestand ein Jahr vor Aufnahme der Schweinebeweidung und die Entwicklung der Brutbestände in den drei Jahren nach Einrichtung der Schweineweide dargestellt. 1998 wurden insgesamt 5 Arten mit insgesamt 9 Brutpaaren bezogen auf eine von den Zielarten besiedelbare Fläche von 9 ha festgestellt. Ein herausragendes Ereignis stellt die Brut eines Wiesenweihenpaares in 1998 im Echeler Bruch dar. Es war die erste dokumentierte Brut dort – leider handelte es sich aber um ein einmaliges Ereignis. Die Revierdichte der berücksichtigten Arten lag bei 1 Revier/ha. Im ersten Jahr nach Einrichtung der Schweineweide steigt die Artenzahl bezogen auf die Gesamtfläche von 9 ha auf 7 Arten an, die Siedlungsdichte (Brutreviere/ha) nimmt im Vergleich zur Ausgangssituation mit 2,79 Revieren/ha auf den Brachen und Röhrichten außerhalb der Schweineweide und 5,25 Revieren/ha im Bereich der Schweineweide deutlich zu. In den beiden Folgejahren sinkt die Zahl der Brutvögel jedoch wieder auf 6 Zielarten im Gesamtgebiet ab, und auch die Revierdichte geht wieder zurück, liegt aber weiterhin deutlich über dem Ausgangszustand (vgl. Tab. 1).
| Tab. 1: | Entwicklung der Brutvogelbestände wertbestimmender Arten in den Eggelwiesen im Zeitraum von 1998 bis 2002. |
Einen Vergleich mit den Verhältnissen in den beiden benachbarten Niedermooren bietet Tab. 2. Dort sind für das Jahr 2002, aus dem von allen drei Flächen nach vergleichbarer Methode erhobene Daten vorliegen, die Brutvogelarten, die Anzahl der Reviere (jeweils linke Spalte) und die Revierdichten (jeweils rechte Spalte, bezogen auf die jeweils für die Zielarten besiedelba-
103
| Tab. 2: | Vergleich der Artenzahlen, Zahl der Brutreviere und Siedlungsdichten (Rev./ha) in den drei Feuchtgebieten im Jahr 2002 (nähere Erläuterungen im Text). |
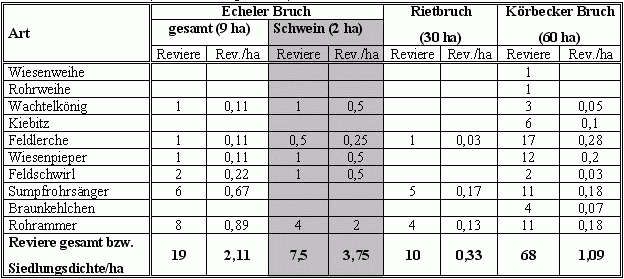
Hinsichtlich der Artenzahl und der Anzahl der Reviere nimmt der Echeler Bruch mit seinen 9 ha eine Stellung zwischen den beiden wesentlich größeren Gebieten (RB: 30 ha, KB: 60 ha) ein. Im Körbecker Bruch waren mit 10 Arten und 68 Revieren mit Abstand die meisten Zielarten und größten Brutbestände anzutreffen. Im Echeler Bruch wurden 2002 6 und im Rietbruch 3 Zielarten mit 19 bzw. 10 Revieren angetroffen. Betrachtet man die Siedlungsdichte, also die Zahl der Reviere pro ha, führt dagegen der Echeler Bruch die Rangliste mit 2,11 Revieren/ha im Vergleich zu 1,09 bzw. 0,33 Revieren an (vgl. Tab. 2). Die höchste Dichte wird mit 3,75 Rev./ha auf der Schweineweide selbst erreicht.
3.1.2 Rastvögel
Die wertbestimmenden Rastvögel in den drei Feuchtgebieten sind mit einigen wenigen Ausnahmen wie Sumpfohreule oder Kranich allesamt der Gruppe der Watvögel (Limikolen) zuzuordnen. Regelmäßig sind während des Zuges Bekassine, Zwergschnepfe, Waldwasserläufer und Bruchwasserläufer anzutreffen. Eine besondere Bedeutung kommt den Flächen während des Frühjahrszuges zu. In Tab. 3 sind die jeweiligen Maximalzahlen für die Bekassine getrennt nach Herbst und Frühjahr für die verschiedenen Gebiete dargestellt. Zu beachten ist, dass für Rietbruch und Körbecker Bruch nicht aus allen Jahren Daten vorliegen.
104
| Tab. 3: | Maximale Anzahl rastender Bekassinen im Echeler Bruch im Zeitraum von 1998 bis 2002, im Vergleich zum Rietbruch und Körbecker Bruch aus den Jahren 2001 und 2002. |
| Echeler Bruch | Rietbruch | Körbecker Bruch | ||||
|---|---|---|---|---|---|---|
| Frühjahr | Herbst | Frühjahr | Herbst | Frühjahr | Herbst | |
|
1998 |
6 | 3 | ||||
|
1999 |
8 | 4 | 5 | - | ||
|
2000 |
69 | 7 | ||||
|
2001 |
26 | 6 | 7 | - | 12 | 5 |
|
2002 |
18 | 7 | 4 | 1 | 8 | 5 |
Während bis zur Einrichtung der Schweineweide die maximale Zahl rastender Bekassinen im Echeler Bruch zwischen 6 und 8 Tieren im Frühjahr und bei maximal 4 Tieren im Herbst lag, nimmt die Zahl in der Folge sprunghaft zu, um dann wieder auf ein relativ hohes Niveau von 18 bis 26 Tieren abzufallen. Während des Herbstzuges sind zwar auch Zunahmen festzustellen – sie erreichen aber nicht die Höhe während des Frühjahrszugs. Die Zahlen aus den wesentlich größeren Vergleichsgebieten sind hingegen deutlich geringer.
Bei den anderen auftretenden Limikolenarten konnten entsprechende Zunahmen nicht festgestellt werden. Sie treten in allen drei Gebieten immer nur in wenigen Individuen auf.
3.2 Laufkäfer
3.2.1 Artengemeinschaft
In den drei Untersuchungsjahren konnten im Echeler Bruch auf der Schweineweide und der angrenzenden Referenzfläche 2.217 Laufkäfer aus 47 Arten nachgewiesen werden. Während der einjährigen Beprobung des Rietbruchs wurden 92 Käfer aus 13 Arten gefangen. Die reinen Fangergebnisse, unabhängig von unterschiedlichen Expositionszeiten und Fallenanzahl sind in Tab. 4 dargestellt. Zusätzlich sind die Daten für den Körbecker Bruch (Kroker 1983) in der Tabelle enthalten.
| Tab. 4: | Aktivitätsdichte der Laufkäfer im Echeler Bruch in den Jahren 2000-02. Zusätzlich sind die Nachweise aus dem Rietbruch (RB) im Jahr 2001 und dem Körbecker Bruch (KB) nach (KROKER 1983) angegeben. |
| Abk.: | BN = Brache nass, SN = Schweineweide nass, BF = Brache frisch, SF = Schweineweide frisch. RL D = Rote Liste Deutschland (TRAUTNER et al. 1997), RL NRW = Nordrhein-Westfalen (SCHÜLE & TERLUTTER 1998); 2 = stark gefährdet, 3 = gefährdet, V = Art der Vorwarnliste, V* = Gefährdungssituation regional stark unterschiedlich; Öko = ökol. Ansprüche nach (BARNDT et al. 1991), (h)/h = (eingeschränkt) hygrophil, hw = hygrophile Waldart, (x) = eingeschränkt xerophil, eu = euryök. |
| Art | RL D | RL NRW | Öko | BN | SN | BF | SF | Summe | RB | KB |
|---|---|---|---|---|---|---|---|---|---|---|
|
Amara communis |
+ |
+ |
(h) |
1 |
7 |
23 |
272 |
303 |
• |
• |
|
Amara lunicollis |
+ |
+ |
(x)(w) |
|
12 |
70 |
204 |
286 |
|
|
|
Bembidion gilvipes |
V* |
3 |
h(w) |
8 |
83 |
28 |
163 |
282 |
• |
• |
|
Agonum afrum/viduum |
+ |
+ |
h |
67 |
140 |
|
10 |
217 |
• /• |
|
|
Bembidion biguttatum |
+ |
+ |
h |
48 |
63 |
2 |
50 |
163 |
|
|
|
Poecilusversicolor |
+ |
+ |
(h) |
|
2 |
39 |
112 |
153 |
• |
|
|
Pterostichus strenuus |
+ |
+ |
(h)w |
20 |
65 |
9 |
24 |
118 |
• |
• |
|
Trechus obtusus / quadristriatus |
+ |
+ |
- |
1 |
62 |
15 |
11 |
89 |
|
• |
105
| Art | RL D | RL NRW | Öko | BN | SN | BF | SF | Summe | RB | KB |
|---|---|---|---|---|---|---|---|---|---|---|
|
Pterostichus diligens |
V |
+ |
h |
30 |
51 |
|
5 |
86 |
• |
• |
|
Epaphius secalis |
+ |
+ |
hw |
|
8 |
36 |
13 |
57 |
|
• |
|
Pterostichus melanarius |
+ |
+ |
eu |
8 |
27 |
6 |
15 |
56 |
• |
|
|
Carabus granulatus |
+ |
+ |
h(w) |
9 |
27 |
|
19 |
55 |
• |
• |
|
Clivina fossor |
+ |
+ |
eu |
6 |
18 |
3 |
28 |
55 |
• |
• |
|
Pterostichus nigrita |
+ |
+ |
h(w) |
5 |
35 |
|
11 |
51 |
• |
• |
|
Harpalus latus |
+ |
+ |
(h)(w) |
|
|
18 |
14 |
32 |
|
|
|
Notiophilus palustris |
+ |
+ |
(h)(w) |
3 |
3 |
8 |
12 |
26 |
|
|
|
Oodes helopioides |
+ |
V |
h |
3 |
20 |
|
1 |
24 |
|
|
|
Bembidion mannerheimii |
+ |
+ |
h(w) |
6 |
13 |
|
1 |
20 |
|
• |
|
Poecilus cupreus |
+ |
+ |
(h) |
|
|
1 |
17 |
18 |
• |
|
|
Carabus nemoralis |
+ |
+ |
(h)(w) |
|
|
2 |
13 |
15 |
|
|
|
Stenolophus teutonus |
+ |
+ |
h |
|
11 |
2 |
1 |
14 |
|
|
|
Bembidion obtusum |
+ |
+ |
(x) |
5 |
|
|
6 |
11 |
|
|
|
Badister lacertosus |
+ |
+ |
(h)w |
|
2 |
4 |
4 |
10 |
|
|
|
Panagaeus cruxmajor |
V |
+ |
h |
|
2 |
2 |
5 |
9 |
|
|
|
Anisodactylus binotatus |
+ |
+ |
(h)(w) |
|
7 |
|
1 |
8 |
|
|
|
Pterostichus vernalis |
+ |
+ |
h |
|
3 |
|
4 |
7 |
|
• |
|
Pterostichus anthracinus |
+ |
+ |
h(w) |
|
5 |
1 |
|
6 |
|
|
|
Acupalpus flavicollis |
+ |
+ |
h |
|
5 |
|
|
5 |
|
|
|
Agonum fuliginosum |
+ |
+ |
h(w) |
1 |
3 |
|
1 |
5 |
|
• |
|
Pterostichus minor |
+ |
+ |
h(w) |
|
4 |
|
1 |
5 |
|
• |
|
Bembidion properans |
+ |
+ |
(x) |
2 |
|
2 |
|
4 |
|
|
|
Harpalus affinis |
+ |
+ |
(x) |
|
|
3 |
1 |
4 |
|
|
|
Amara convexior |
+ |
+ |
(x) |
|
|
3 |
|
3 |
|
|
|
Amara plebeja |
+ |
+ |
eu |
|
1 |
|
2 |
3 |
|
|
|
Amara similata |
+ |
+ |
eu |
2 |
|
|
|
2 |
|
|
|
Bembidion guttula |
V* |
+ |
h |
|
|
|
2 |
2 |
|
|
|
Elaphrus uliginosus |
2 |
2 |
h |
|
2 |
|
|
2 |
|
|
|
Leistus ferrugineus |
+ |
+ |
(x)(w) |
|
|
1 |
1 |
2 |
|
• |
|
Stenolophus mixtus |
+ |
+ |
h |
1 |
1 |
|
|
2 |
|
|
|
Trichocellus placidus |
+ |
+ |
hw |
1 |
1 |
|
|
2 |
|
|
|
Bembidion lampros |
+ |
+ |
(x)(w) |
|
|
1 |
|
1 |
|
|
|
Bembidion tetracolum |
+ |
+ |
eu |
|
|
|
1 |
1 |
|
|
|
Calathus melanocephalus |
+ |
+ |
(x) |
|
|
|
1 |
1 |
|
|
|
Leistus terminatus |
+ |
+ |
h(w) |
|
|
|
1 |
1 |
|
• |
|
Ophonus rufibarbis |
+ |
+ |
(x)(w) |
|
|
1 |
|
1 |
|
|
|
Loricera pilicornis |
+ |
+ |
(h) |
|
|
|
|
|
|
• |
|
Dyschirius globosus |
+ |
+ |
(h) |
|
|
|
|
|
|
• |
|
Synuchus vivalis |
+ |
+ |
(x) |
|
|
|
|
|
|
• |
|
Agonum thoreyi |
+ |
+ |
h |
|
|
|
|
|
|
• |
|
Amara aulica |
+ |
+ |
eu |
|
|
|
|
|
|
• |
|
Chlaenius nigricornis |
V* |
V |
h |
|
|
|
|
|
• |
|
|
Individuen |
|
|
|
227 |
683 |
280 |
1027 |
2217 |
92 |
499 |
|
Arten |
|
|
|
21 |
31 |
25 |
36 |
47 |
13 |
20 |
|
Aktivität |
|
|
|
50,6 |
79,5 |
56,4 |
123,3 |
|
|
|
106
Dominiert wird die Artengemeinschaft zum einen von typischen Arten des Feuchtgrünlands wie z. B. Agonum afrum/viduum, Bembidion gilvipes, B. biguttatum, Epaphius secalis, Pterostichus diligens und Pt. nigrita. Zum anderen sind einige hinsichtlich der Bodenfeuchte eher anspruchslose und in der Kulturlandschaft weit verbreitete Arten auf den frisch-feuchten Teilflächen häufig: hierzu gehören Amara communis, A. lunicollis, Poecilus versicolor, Pterostichus melanarius, Harpalus latus und Clivina fossor. Zwar ebenfalls feuchteliebend, aber trotzdem mit einer weiteren ökologischen Amplitude ausgestattet, sind Carabus granulatus oder Pterostichus strenuus.
Es zeichnet sich deutlich ab, dass der standörtliche Feuchtegradient erheblichen Einfluss auf die Taxozönosen im Echeler Bruch ausübt. Viele Arten sind mit ihren Vorkommen auf die nassen bzw. frisch-feuchten Bereiche beschränkt (s. auch Tab. 5). Es kann jedoch ebenfalls festgehalten werden, dass auf der Schweineweide jeweils deutlich mehr Arten nachgewiesen wurden als auf der benachbarten Brachfläche. Gleichzeitig ist auf der Schweineweide die Aktivität der Laufkäfer höher: auf den trockeneren Flächen z. B. mehr als doppelt so hoch wie auf vergleichbaren Flächen der Brache.
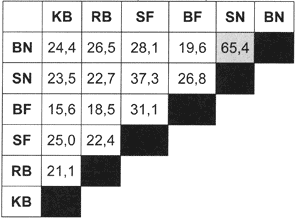
In Tab. 2 ist die Artenidentität zwischen den drei betrachteten Niedermooren aufgetragen. Auffällig sind die überwiegend geringen Übereinstimmungen.
| Tab. 5: | Artenähnlichkeit nach SÖRENSEN der drei betrachteten Niedermoorstandorte unter Berücksichtigung der 4 Teilflächen im Echeler Bruch |
| Abk.: | BN = Brache nass, SN = Schweineweide nass, BF = Brache frisch, SF = Schweineweide frisch, KB = Körbecker Bruch, RB = Rietbruch |
3.2.2 Landnutzung
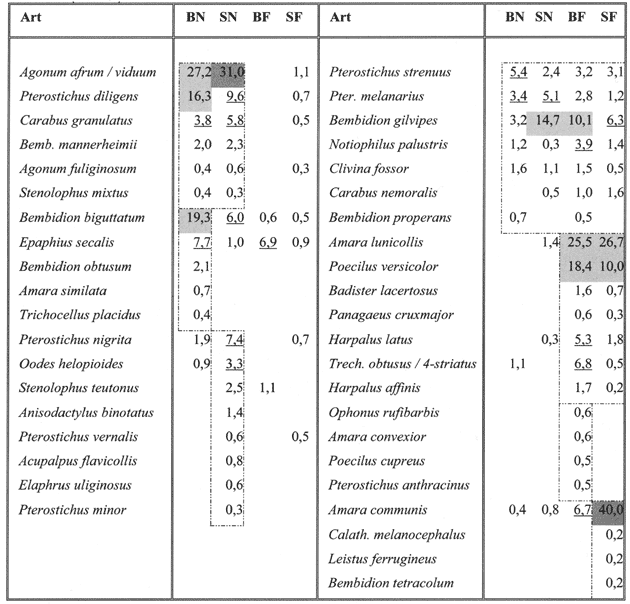
Für den weitergehenden Vergleich der Auswirkungen der Schweinebeweidung wurde die Anzahl der verwendeten Fallen angeglichen (s. Kap. 2) und die Aktivitätsdichte der Laufkäfer auf 100 Fallentage standardisiert. Auf diese Weise werden im Folgenden noch 1.223 Individuen aus 42 Arten berücksichtigt. Die Aktivitätsdominanz für den Echeler Bruch ist in Tab. 6 dargestellt.
Zunächst fällt wiederum auf, dass der Feuchtegradient der Untersuchungsfläche sich sehr deutlich in den Laufkäfer-Zönosen widerspiegelt (vgl. auch Abb. 2). Während auf den sumpfig-nassen Teilbereichen Pter. diligens, Agonum afrum und A. viduum dominieren, erreichen auf den frisch-feuchten Flächen Poecilus versicolor und Amara lunicollis die höchste Aktivitätsdominanz. Weiterhin wurden in den nassen Flächen Carabus granulatus, Stenolophus mixtus, Agonum fuliginosum und Bemb. mannerheimi häufiger gefangen – im Frisch-feuchten Badister lacertosus und Panagaeus cruxmajor. Demgegenüber gibt es einige euryöke Offenlandsarten ohne erkennbare Präferenzen für Feuchte und Nutzung der Flächen, wie Pter. melanarius, Pter. strenuus, Bemb. properans, Carabus nemoralis, Clivina fossor und Notiophilus palustris.
Einige Arten zeigen auf der Schweineweide eine deutlich höhere Aktivität: Oodes helopioides, Pter. nigrita, Stenolophus teutonus, Elaphrus uliginosus, Pter. minor (alle im Nassen) sowie Amara communis (im frischen Bereich der Weide). Umgekehrt können auch „Präferenzen“ für die Brache beobachtet werden. Im Einzelnen bei Bemb. biguttatum und Epaphius secalis (Nass) und Harpalus latus, Harp. affinis und Trechus ssp. (frisch-feucht).
107
| Tabelle 1: | Aktivitätsdominanz der Laufkäfer am Standort Echeler Bruch – getrennt nach Nutzung und Bodenfeuchte (vgl. Tab. 1). (Eu)dominante Arten (> 30 bzw. 10 %) sind (dunkel)grau, subdominante Arten (> 3,2 %) sind unterstrichen. |
3.2.3 Funktionelle Merkmale
Die Bedeutung ausgewählter funktioneller Merkmale – also taxa-bezogenen Merkmalen, die auf Umweltfaktoren reagieren – ist bei Laufkäfern durch die Arbeiten von LARSSON (1939) und LINDROTH (1949) seit langem belegt. In jüngerer Zeit wurden das Analysespektrum um weitere Merkmalsausprägungen erweitert (z. B. RIBERA et al. 2001; BLAKE et al. 2003).
1. Ökologische Anspruchstypen
Der charakteristische Feuchtegradient im Untersuchungsgebiet Echeler Bruch wird auch bei der Betrachtung der ökologischen Ansprüche der nachgewiesenen Laufkäferindividuen deutlich. Während im nassen Teilbereich über 50 % der Laufkäfer den streng hygrophilen Arten zuzurechnen ist, liegt deren Anteil im frisch-feuchten nur bei max. 10 %. Dort ist dafür der Anteil an leicht hygrophilen Laufkäfern sowie an leicht xerophilen Waldarten deutlich größer.
Allerdings sind auch unterschiedliche Entwicklungen als Ergebnis der Nutzungsunterschiede im Verlauf der drei Jahre zu erkennen (Abb. 3). Während sich auf der nassen Brache (BN) kaum Änderungen ergeben, ist im Bereich des nassen Teils der Schweineweide (SN) eine Zunahme der hygrophilen und euryöken Arten auf Kosten der Feuchtwaldarten festzustellen. Ganz anders
108
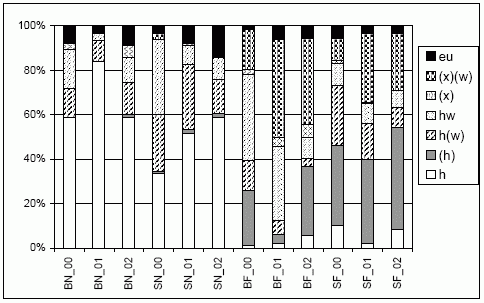
| Abb. 3: | Verteilung der ökologischen Anspruchstypen der Laufkäferindividuen (nach u. a. Barndt et al. 1991; Raths & Riecken 1999; Turin 2000) getrennt nach Jahr (2000 bis 2002) und Standort (BN, SN, BF, BN) im UG Echeler Bruch. |
| Abk.: | BN/F - Brache nass/frisch-feucht, SN/F - Schweineweide nass/frisch-feucht, eu - euryök, x - xerophil, h - hygrophil, w - Waldart |
2. Flugdynamischer Typ
Entsprechend der standörtlichen Differenzierung herrschte bereits zu Beginn der Untersuchung ein deutlicher Unterschied zwischen den nassen und den frisch-feuchten Teilbereichen – letztere beherbergen einen wesentlich höheren Anteil an konstant langflügeligen Laufkäfern (> 50 %).
Dort steigt deren Anteil im Verlauf der Untersuchung sowohl auf Schweineweide als auch auf der Brache auf ca. 80 % (Abb. 4). Auf der nassen Brache nimmt hingegen der Anteil an Arten mit dimorpher Flügelausbildung zu (ebenfalls ca. 80 %) – im nassen Bereich der Schweineweide (SN) ergeben sich kaum Veränderungen.
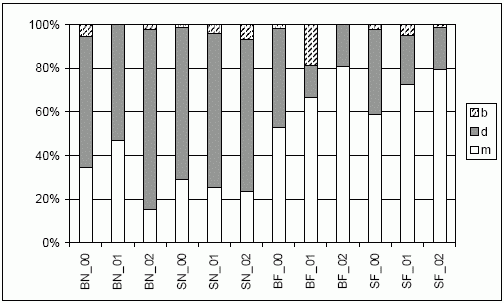
| Abb. 4: | Verteilung der Flügelausbildung der Laufkäferindividuen (nach u.a. BARNDT et al. 1991; RATHS & RIECKEN 1999; TURIN 2000) getrennt nach Jahr (00 bis 02) und Standort im Echeler Bruch. |
| Abk.: | BN/F - Brache nass/frisch-feucht, SN/F - Schweineweide nass/frisch-feucht; b = brachypter (kurzflügelig), d = dimorph, m = makropter (langflügelig) |
109
3. Entwicklungstypen
Insgesamt dominieren im Echeler Bruch so genannte „Frühjahrsarten“, d.h. Laufkäfer mit einer Reproduktion im Frühsommer und einer Imaginalüberwinterung (vgl. LARSSON 1939). Bezieht man die Arten, die einen aktiven Herbstbestand aufweisen, mit ein, so stellen sie zu Beginn der Untersuchungen zwischen 70 und 95 % der Individuen (Abb. 5). Nach Ablauf der drei Untersuchungsjahre hat sich diese Tendenz weiter angeglichen (85-95 %), wobei die Entwicklungen im Einzelnen etwas unterschiedlich verlaufen. Insbesondere die Herbstarten, die auf Brache frisch (BF) und Schweineweide nass (SN) zu Beginn der Untersuchung noch in etwas größerer Zahl auftreten, nehmen deutlich ab. Gleichzeitig ist auf den nassen Teilflächen die Zunahme von Arten mit instabilem Entwicklungstyp zu beobachten.
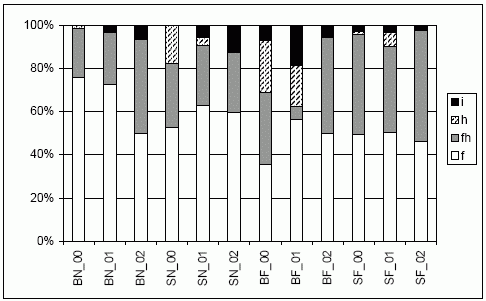
| Abb. 5: | Verteilung der Entwicklungstypen der Laufkäferindividuen (nach u.a. Barndt et al. 1991; Raths & Riecken 1999; Turin 2000) getrennt nach Jahr (00 bis 02) und Standort im Echeler Bruch. |
| Abk.: | BN/F - Brache nass/frisch-feucht, SN/F - Schweineweide nass/frisch-feucht; f = Frühjahrsart, fh = Frühjahrsart mit aktivem Herbstbestand, h = Herbstart, i = instabiler Entwicklungstyp. |
4. Aktivitätsrhythmik
Die zu Tage tretenden Unterschiede folgen in diesem Fall nicht dem standörtlichen Gradienten. Zu Beginn der Untersuchung nehmen die tagaktiven Arten auf Brache nass (BN) und Schweineweide frisch (SF) einen besonders großen Anteil von ca. 80 % ein, wobei die folgende Entwicklung gegenläufig ist. Während auf der Brache der Anteil auf die Hälfte zurückgeht, steigt er auf der SF auf über 90 %. Kaum Veränderungen ergeben sich auf der nassen Schweineweide – hier halten sich beide Aktivitätstypen in etwa die Waage. Demgegenüber nimmt der Anteil der tagaktiven Laufkäfer auf der trockeneren Brache deutlich zu – von 60 auf ca. 80 %.
110
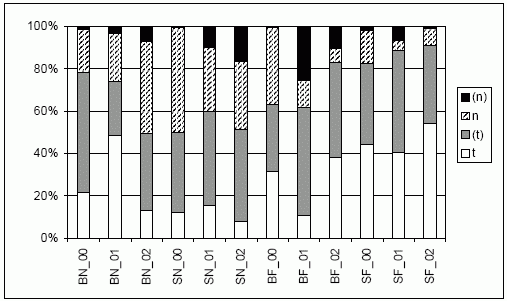
| Abb. 6: | Verteilung der Aktivitätsrhythmik der Laufkäferindividuen (nach u.a. Kegel 1990; Barndt et al. 1991; Raths & Riecken 1999; Turin 2000) getrennt nach Jahr (00 bis 02) und Standort im Echeler Bruch (für 98 Ind. erfolgte keine Einstufung – sie bleiben in der Darstellung unberücksichtigt). |
| Abk.: | BN/F - Brache nass/frisch-feucht, SN/F - Schweineweide nass/frisch-feucht; (t)/t = (überwiegend) tagaktiv, (n)/n = (überwiegend) nachtaktiv. |
3.3 Abiotik
Die verschiedenen Teiluntersuchungsflächen unterscheiden sich signifikant hinsichtlich ausgewählter abiotischer Parameter. Es zeigt sich, dass bspw. der Rohboden auf den Schweineweiden signifikant größere Flächenanteile einnimmt (H-Test; n = 202; df = 3; Chi² = 46,3; p < 0,001) – dies ist besonders auf der nassen Teilfläche zu erkennen (Abb. 7). Bzgl. der Vegetationshöhe bestehen die Unterschiede in erster Linie zwischen den verschiedenen Bodenfeuchtestufen des Standortes (ANOVA; n = 132; df = 3; F = 7,6; p < 0,001). Auf den frisch-feuchten Bereichen waren im Mittel höhere Pflanzenbestände anzutreffen. Die bevorzugt bewühlten nassen Bereiche der Schweineweide weisen signifikant niedrigere Vegetationshöhen auf als alle anderen Teilflächen (LSD Post-hoc Test).
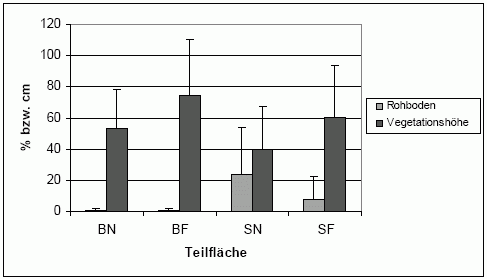
| Abb. 7: | Kenngrößen abiotischer Parameter im jeweiligen Fallenumfeld (1 m², Mittelwert der 3 Untersuchungsjahre incl. Standardabweichung) auf den Teiluntersuchungsflächen im Echeler Bruch. |
| Abk.: | BN/F - Brache nass/frisch-feucht, SN/F - Schweineweide nass/frisch-feucht. |
111
4 Diskussion
4.1 Avifauna
Die aufgezeigten Entwicklungen im Echeler Bruch nach Einrichtung der Schweineweide verdeutlichen, dass die neu etablierte Nutzungsform aus Naturschutzsicht deutliche Vorteile gegenüber dem Ausgangszustand "Brache" mit sich bringt. Dies macht sich nicht so sehr in einer Zunahme der Artenzahlen als vielmehr in einer deutlichen Erhöhung der Siedlungsdichten bei den Wiesen- und Röhrichtbrütern bemerkbar. Diese positive Entwicklung ist nicht nur auf die Schweineweide beschränkt, vielmehr scheint die mit 2 ha recht kleinflächige Weide auch den Brutvogelbestand auf der verbleibenden, wesentlich größeren Restfläche positiv zu beeinflussen. Da aufgrund der Datenlage nur das Jahr 1998 als Vergleich herangezogen werden konnte, wurden zur Absicherung der Ergebnisse die beiden benachbarten Bruchflächen mit in die Untersuchungen bzw. Auswertungen einbezogen. Auch dieser Vergleich deutet in die gleiche Richtung: Die Siedlungsdichten liegen im Echeler Bruch in seiner Gesamtheit und insbesondere auf der Schweineweide doppelt bis 3,5mal so hoch wie im Körbecker Bruch. Im Vergleich zum Rietbruch ist die Siedlungsdichte auf der Schweineweide sogar 10mal höher. Dieses für den Rietbruch ungünstige Ergebnis dürfte v. a. auf die dortige gute Strukturierung durch Bäume und Gehölze zurückzuführen sein, die sich auf eine Besiedlung durch die Zielarten (Wiesen, Röhrichtbrüter) negativ auswirkt. Allerdings wurde schon bei der Aufbereitung der Ergebnisse versucht, diese Effekte möglichst nicht einfließen zu lassen, indem die Siedlungsdichten nur für die geeigneten Flächen berechnet wurden.
Erklärbar sind die deutlichen Unterschiede mit den Habitat- und Strukturansprüchen der charakteristischen Brutvögel des feucht-nassen Offenlandes. Arten wie die Feldlerche benötigen niedrigwüchsige oder lückige Vegetationsbestände, wie sie auf den stärker bewühlten Bereichen anzutreffen sind. Wiesenpieper oder Braunkehlchen bevorzugen dagegen eine gut strukturierte Krautschicht mit kurzrasigen und deckungsreichen Bereichen sowie höhere Strukturen, die sich als Sing- bzw. Sitzwarten eignen. Auch diese gehören zur charakteristischen Ausstattung einer Schweineweide, insbesondere die umfangreichen Zäune, aber auch die Schweinehütten, stellen hervorragende Singwarten dar. Da durch das selektive Weideverhalten der Schweine großflächig Hochstauden und Röhrichtbestände bestehen bleiben, findet sich ein zusätzlicher Strukturtyp auf den Flächen, der von entsprechenden Arten (Rohrammer, Feldschwirl oder Sumpfrohrsänger) als Lebensraum genutzt wird. Dieses vielfältige Struktur- und Habitatangebot scheint auch den Ansprüchen des europaweit gefährdeten Wachtelkönigs entgegenzukommen, der im Echeler Bruch schon seit längerem als Durchzügler bekannt war, seit Errichtung der Schweineweide aber zum Brutvogel geworden ist.

| Abb. 8: | Echeler Bruch (Foto: Frank Grawe) |
Auch für rastende Bekassinen sind Schweineweiden in Feuchtgebieten von hoher Attraktivität, wie durch die Bestandsentwicklungen im Echeler Bruch belegt wird. Für diese Art sind es die im Spätsommer/Herbst durch die Schweine geschaffenen großflächigen Schlammflächen, die das benötigte stocherfähige Substrat für die Nahrungssuche bereitstellen.
Vergleichbare Ergebnisse hinsichtlich Brut- und Rastvögeln wurden im Rahmen des Forschungsvorhabens auch auf Schweineweiden im Auengrünland des Elbtals bei Lenzen erzielt (BEINLICH et al., in Vorb.).
112
4.2 Laufkäfer
4.2.1 Artengemeinschaft
Die Artenzusammensetzung der Taxozönosen im Echeler Bruch wird in erster Linie von der standörtlichen Heterogenität der Bodenfeuchte bestimmt (vgl. Abb. 3). Erst in zweiter Linie spielt auch die Nutzung der Flächen für die weitere Differenzierung eine Rolle. Dies deckt sich mit den detaillierten Untersuchungen in verschiedenen Niedermoorgebieten Norddeutschlands – auch dort erwiesen sich beide Faktoren als entscheidend (FISCHER et al. 1998; IRMLER et al. 1998). FUELLHAAS (1998) betont hingegen in erster Linie die Bedeutung des Vernässungsgrads bzw. der Dauer der Überstauung für die Herausbildung spezifischer Artenzusammensetzungen. In den von ihm untersuchten Niedermoorflächen spielte die Bewirtschaftungsintensität, d. h. das Mahdregime, nur eine untergeordnete Rolle, da es direkt von der Befahrbarkeit der Flächen und somit vom Vernässungsgrad abhing. Auf einer anderen extensiv genutzten Schweineweide zeigte sich, dass die Ausprägung der Laufkäfergemeinschaft durchaus auch vom Zeitpunkt der Beweidung bestimmt werden kann (HILL et al. 2003).

| Abb. 9: | Düppeler Weideschweine im Echeler Bruch (Foto: Frank Grawe) |
Die im Echeler Bruch festgestellten Artenzahlen (zwischen 21 und 35 pro Fläche) – entsprechen Befunden aus ähnlichen Lebensräumen (Feuchtgrünland und -brachen, Schilfgebieten), (vgl. DÜLGE et al. 1994; HANDKE 1995; FRITZE 1998; MENKE 2000). Es konnten durchgehend mehr Laufkäferarten auf den Schweineweiden nachgewiesen werden. Dies bestätigt eine der Ausgangshypothesen, dass die gegenüber Brachen durch Wühltätigkeit und selektives Fraßverhalten abwechslungsreicher strukturierten Schweineflächen (z. B. Abb. 7) eine höhere Artenvielfalt aufweisen. Gerade in den für Laufkäfer entscheidenden Parametern Bodenfeuchte und Mikroklima ist hier ein größeres Angebot an Nischen vorhanden. Auch in allen weiteren Untersuchungen zur extensiven Schweinebeweidung (überwiegend im Rahmen dieses Projekts) hat sich dieser Zusammenhang bestätigt (HEROLD 2001; HILL et al. 2003; HILL et al. 2004). Dass die Bedeutung extensiver Beweidungssysteme für eine hohe Artenvielfalt bei Laufkäfern hierbei nicht auf Schweine begrenzt ist, sondern auch für andere Weidetiere zutrifft, zeigen z. B. SCHULZ (2002) oder LEDERBOGEN et al. (2004).
Die hohe Aktivitätsdichte auf der Schweineweide im Vergleich zur Brache dürfte auf den geringeren Raumwiderstand (vgl. HEYDEMANN 1957) zurückzuführen sein. In den durch Fraß, Wühltätigkeit und Tritt aufgelockerten Bereichen ist demzufolge ein schnelleres Fortbewegen der Käfer möglich, was eine erhöhte Fangwahrscheinlichkeit bedingt.
In den nassen Bereichen findet sich eine typische Zönose für Niedermoore, die von Kennarten des Feuchtgrünlands: Pt. diligens, Pt. nigrita, Pt. minor, Agon. afrum/viduum, Car. granulatus, Epaphius secalis, Oodes helopioides, Sten. mixtus und der Niedermoore: Agon. fuliginosum, Sten. teutonus dominiert wird (vgl. TIETZE 1973; DÜLGE et al. 1994; FISCHER et al. 1998). Einige Arten erreichen auf der Schweineweide deutlich höhere Aktivitätsdominanzen.
Als anspruchsvoll gilt Oodes helopioides, der als eine der wenigen Laufkäfer auch dauerhaft überstaute Flächen zu besiedeln vermag. In Westfalen ist er aus allen Landesteilen bekannt – in der Ebene aber deutlich häufiger (A&SZLIG;MANN & STARKE 1990). Bundesweit stark gefährdet und in Westfalen generell selten ist Elaphrus uliginosus (RUDOLPH 1976). Als optischer Jäger ist er
113
Das Fehlen weiterer stenöker Niedermoorarten ist zum einen wahrscheinlich eine Folge der Entwässerung und landwirtschaftlichen Nutzung der Flächen in der Vergangenheit. Diese „Epoche“ konnten nur anpassungsfähige Arten überdauern. Eine schnelle Wiederbesiedlung aus dem Umfeld ist aufgrund der isolierten Lage und dem weitgehenden Fehlen von möglichen Spenderpopulationen (s. Artenbestand RB bzw. KB, Tab. 1) nur sehr eingeschränkt möglich. Zum anderen wird auch in größeren, weitgehend intakten Niedermoorgebieten Norddeutschlands der überwiegende Teil der Laufkäferzönose von (Feucht)Grünlandarten gestellt (DÜLGE et al. 1994; FISCHER et al. 1998).
Die höher gelegenen Teilbereiche mit schlechterer Wasserversorgung zeigen dagegen deutlich mehr Übereinstimmung mit Zönosen des mesophilen Grünlands. Es dominieren eurytope Grünlandarten wie Amara communis, A. lunicollis und Poec. versicolor. Zudem dringen auch trockenheitsliebende Ackerbewohner, z.B. Bembidion lampros, Harpalus affinis, Ophonus rufibarbis, Calathus melanocephalus und Amara convexior aus dem Umfeld ein. Dieser Umstand ist auch von anderen Niedermooren mit gestörtem Wasserhaushalt und landwirtschaftlicher Nutzung beschrieben (z. B. FISCHER et al. 1998; FUELLHAAS 1998). Insofern ist allein eine hohe Artenzahl nicht als wertbestimmendes Kriterium für Niedermoore zu werten.
Vergleich mit den anderen Niedermoorflächen
Die Artenübereinstimmung zwischen Echeler Bruch, Rietbruch und Körbecker Bruch bzw. den verschiedenen Teilflächen des Echeler Bruch ist generell sehr niedrig (vgl. Tab. 2). Die einzige größere Ähnlichkeit besteht zwischen den nassen Teilflächen im Echeler Bruch (BN und SN). Weiterhin sind gewisse Ähnlichkeiten zwischen den beiden Teilen der Schweineweide zu konstatieren. Sowohl Rietbruch als auch Körbecker Bruch besitzen kaum Gemeinsamkeiten untereinander bzw. mit dem Echeler Bruch. Insgesamt ist aber insbesondere im Rietbruch die Zahl an nachgewiesenen Arten als sehr gering einzustufen. Auch hier treten zwar Feuchtgebietsbewohner wie Pterostichus diligens, Pter. nigrita und Agonum afrum/viduum auf, die anspruchslosen Offenlandsarten (Poecilus cupreus/versicolor, Pter. melanarius) nehmen aber einen breiteren Raum ein. Bereits MÜLLER et al. (1990) führen in ihrer Übersichtskartierung im Rietbruch mit Ausnahme von P. cupreus keine biotoptypischen Arten auf, so dass der von uns gewonnene Eindruck bestätigt wird. Ob die Artenarmut auf die bis vor wenigen Jahren noch praktizierte intensive Grünlandwirtschaft zurückzuführen ist, ist unklar.
Im Körbecker Bruch hingegen liegt der Anteil an Feuchtgrünlandskennarten (Stand 1983) deutlich höher, wobei auch dort einige typische Arten fehlen, die im Echeler Bruch auftreten (z.B. Bembidion biguttatum, Oodes helopioides). Allerdings ist mit Agonum thoreyi zumindest ein stenöker Röhrichtbewohner belegt, von dem kein Nachweis im Echeler Bruch gelang (ein Vorkommen aufgrund der kletternden Lebensweise aber auch nicht auszuschließen ist). Auch hier könnte die relative Artenarmut ein Ausdruck der in den 1960er und 1970er intensivierten Grünlandnutzung sein, die 1983, dem Jahr der Erhebungen durch KROKER, noch anhielt und erst mit Ausweisung als NSG wieder nachließ (vgl. LOHR 2005, in diesem Heft).
Allerdings werden auch die verschiedenen Erfassungszeiträume und -methodiken (z. B. im Rietbruch nur ein Jahr mit im Vergleich zum Echeler Bruch deutlich reduzierter Fallenzahl) für die beobachteten Unterschiede mit ausschlaggebend sein.
4.2.2 Funktionelle Merkmale
Generell scheint das spezifische Weideverhalten der Schweine mit ihrem ausgesprochenen Drang zum Wühlen in Feuchtgebieten verschiedene ökologische Anspruchstypen auf derselben Fläche zu begünstigen. So konnte bereits mehrfach gezeigt werden, dass in Bach- und Flussauen
114
Grundsätzlich sind diese Effekte auch im Echeler Bruch zu erkennen (vgl. 3.2) – sie werden allerdings durch die parallel verlaufende Wiedervernässung infolge der verlandenden Gräben z. T. überlagert. In der besonders stark vernässten Teilfläche der Schweineweide (SN) nimmt dementsprechend der Anteil an hygrophilen Offenlandsarten auf Kosten von Feuchtwaldbewohnern zu (vgl. auch HILL et al. 2005). In den durch geringere Wühlaktivität gekennzeichneten frisch-feuchten Bereichen (s. 3.3) sind die Unterschiede zur Brache hingegen sehr gering.
Typischerweise ist in Feuchtgebieten mit hohen Grundwasserständen der Prozentsatz an Larvalüberwinterern („Herbstarten“) deutlich reduziert, da es den Larven schwerer fällt, den für sie ungünstigen Lebensumständen auszuweichen (LARSSON 1939). Dieser Umstand ist auch auf den Untersuchungsflächen zu erkennen – wobei der starke Rückgang der Herbstarten im Bereich der nassen Ausprägung der Schweineweide ins Auge fällt. Die Zunahme an Arten mit instabilem Reproduktionsverhalten könnte dahingehend interpretiert werden, dass die Arten flexibel auf die sich ändernden Grundwasserstände reagieren können. MENKE (2000) fand auf unterschiedlich genutzten Feuchtgrünlandflächen in der Bremer Wesermarsch einen durchschnittlichen Anteil an Larvalüberwinterern von 19 %, bei HANDKE (1995) lag er unter 10 %. Im Echeler Bruch erreichte er im Mittel knapp 7 % und fiel im 3. Jahr auf unter 0,5 %. Ein konsistenter Unterschied je nach Nutzung, wie er von MENKE (2000) festgestellt wurde, trat in unserem Fall nicht auf.
Nach RIECKEN (1992) kann der Anteil an makropteren (langflügeligen) Arten einer Laufkäferzönose als Maß für die Dynamik bzw. Stabilität eines Lebensraums herangezogen werden. So treten bspw. in anthropogen gestörten Nutzsystemen (Intensiv-Äckern), aber auch in von hoher Dynamik geprägten Biotopen (z. B. Fließgewässerufern, Spülfeldern), überwiegend kleine, flugfähige Laufkäfer auf. Auf mit Schweinen beweideten trockenen Ackerbrachen mit hoher raum-zeitlicher Dynamik kommt es ebenfalls zu einer Zunahme von flugfähigen Tieren (HILL et al. 2003). Tatsächlich ist dieser Merkmalstypus auch im Echeler Bruch in den frisch-feuchten Bereichen vorherrschend – die beobachteten Unterschiede werden allerdings nicht von der Flächennutzung, sondern von der Bodenfeuchte bestimmt. Dies ist überraschend, da z. B. MENKE (2000) sehr viel mehr dimorphe Käfer auf ungenutzten Brachen und Röhrichten als auf Wiesen und Weiden finden konnte. Im vorliegenden Fall konzentrieren sie sich unabhängig von der Nutzung auf den nassen Teilflächen. Beiden Untersuchungen gemeinsam ist hingegen der geringe Anteil an brachypteren Arten, denen eine ausweichende Reaktion auf die wechselnden Umweltbedingungen schwerer fallen würde.
Die diurnale Aktivitätsrhythmik von Laufkäfergemeinschaften wird entscheidend von der grundlegenden Lebensraumstruktur bestimmt. So sind in der Kulturlandschaft vermehrt tagaktive Arten (LUFF 1978; KEGEL 1990), im Wald mehr nachtaktive anzutreffen (z.B. LAUTERBACH 1964). Während sich für die frisch-feuchten Teilflächen, die mehr den Charakter eines Agrarbiotops besitzen, diese Beobachtungen bestätigen lassen, scheinen sie für offene Feuchtgebiete nicht ohne weiteres übertragbar. Auf beiden vernässten Flächen fand sich im dritten Jahr ein fast ausgeglichenes Verhältnis von beiden Aktivitätstypen. Dies lässt sich dadurch erklären, dass in erster Linie die hohen Feuchteansprüche für die Nachtaktivität der Arten verantwortlich sind (KEGEL 1990) – in Niedermooren mit hohem Grundwasserstand aber ein Vorkommen auch ohne Beschattung möglich ist.
Gleichzeitig besteht ein Zusammenhang zwischen Entwicklungstyp und Aktivitätsrhythmik der einzelnen Arten: Larvalüberwinterer sind
115
5 Ausblick
Die Untersuchungen zeigen, dass die extensive Schweinebeweidung durchaus in der Lage ist, den typischen Artenbestand der Niedermoore in der Warburger Börde zu sichern. Nach dem aktuellen Kenntnisstand kommt dem Echeler Bruch eine besonders große Bedeutung für den Erhalt der typischen Laufkäfergemeinschaften des Naturraums zu, der von den anderen betrachteten Niedermoorresten in dieser Form nicht geleistet werden kann. Darüber hinaus gibt es einige seltene bzw. gefährdete Vogel- und Laufkäfer-Arten, die – möglicherweise im Zusammenhang mit der Wiedervernässung des Echeler Bruch – von der Beweidung profitieren. Hier sind an erster Stelle der Wachtelkönig (Crex crex), rastende Bekassinen (Gallinago gallinago) und andere Watvögel sowie der Dunkle Uferläufer (Elaphrus uliginosus) zu nennen.

| Abb. 10: | Düppeler Weideschweine (Foto: Frank Grawe) |
Als entscheidende Stellgröße für den Erfolg der Beweidung haben sich der Zeitpunkt und die Intensität (Anzahl der Tiere, Dauer der Beweidung) herauskristallisiert – wobei pauschale Hinweise nicht zielführend sind. Im vorliegenden Fall ist eine saisonale Nutzung der Flächen außerhalb der Hauptbrutzeit der Vögel entscheidend. Die Beweidung im Spätsommer/Herbst ermöglicht eine „intensive“ Wühltätigkeit aufgrund der Nährstoffverhältnisse der Futterpflanzen (Verlagerung in Speicherorgane) und der Bodenfeuchte (bessere „Grabbarkeit“) (vgl. MICKLICH 1996).
Leider bestehen für die extensive Freilandhaltung von Hausschweinen unter den bisherigen EU-Rahmenbedingungen zur Tierhygiene und Agrarförderung eine Vielzahl von Hindernissen (z. B. Zäunung, fehlende Prämien, vgl. HILL et al. 2004). Allerdings entstehen aufgrund der Änderungen in der EU-Agrarpolitik, insbesondere der Einführung einer Grünland-Flächenprämie, neue Rahmenbedingungen. Hiervon könnte ggf. auch die aus Sicht des Tierschutzes und der Landschaftsästhetik wünschenswerte Nutzungsform Schweinefreilandhaltung profitieren – vorausgesetzt, der Verbraucher honoriert die artgerechte Haltung durch die Bereitschaft, auch einen höheren Preis zu bezahlen.
Danksagung
Das Bundesministerium für Bildung und Forschung (BMBF) förderte diese Untersuchungen im Rahmen des Projektes “Schweinefreilandhaltung im Rahmen der Landschaftspflege”, Förder-Kennzeichen 01 LN 0002.
Die NRW-Stiftung Kultur, Heimat und Naturschutz unterstützte das Vorhaben durch die Bereitstellung der Tiere und diverser Weideeinrichtungen.
Die Bezirksregierung Detmold stellte die benötigten Flächen für das Vorhaben im Echeler Bruch zur Verfügung.
Herr Andreas Malten (Dreieich) überprüfte kritische Laufkäfer-Individuen.
Herr Herbert Schröder (Borgentreich) lieferte wertvolle Hinweise zur Vogelwelt der Brüche.
Ihnen allen sei an dieser Stelle herzlich gedankt!
116
Literatur
AßMANN, T. & W. Starke (1990): Coleoptera Westfalica: Familia Carabidae, Subfamiliae Callistinae, Oodinae, Licininae, Badistrinae, Panagaeinae, Colliurinae, Aephidiinae, Lebiinae, Demetriinae, Cymindinae, Dromiinae et Brachininae. – Abhandlungen des Landesmuseums für Naturkunde in Münster / Westfalen 52(1): 3-61.
BARBER, M. S. (1931): Traps for cave-inhabiting Insects. – Journ. Eliska MitchBarber, M Sel Science Soc. 46: 259-266
BARNDT, D., BRASE, S., GLAUCHE, M., GRUTTKE, H., KEGEL, B., PLATEN, R. & WINKELMANN, H. (1991): Die Laufkäferfauna von Berlin (West) - mit Kennzeichnung und Auswertung der verschollenen und gefährdeten Arten (Rote Liste, 3. Fassung). in: Rote Listen der gefährdeten Pflanzen und Tiere in Berlin, Schwerpunkt Berlin (West). AUHAGEN, A., PLATEN, R. & SUKOPP, H. (Hrsg.), Berlin. 6: 243-275.
BEINLICH, B. (1998): Die Schweineweide als dynamisches Element in der Kulturlandschaft. – Schriftenreihe für Landschaftspflege und Naturschutz 56: 317-336.
BIBBY, C. J., N. D. BURGESS & D. A. HILL (1995): Methoden der Feldornithologie: Bestandserfassung in der Praxis. – Radebeul, Neumann.
BLAKE, S., MCCRACKEN, D.I., EYRE, M.D., GARSIDE, A. & FOSTER, G.N. (2003): The relationship between the classification of Scottish ground beetle assemblages (Coleoptera, Carabidae) and the National Vegetation Classification of British plant communities. – Ecography 26(5): 602-616.
CONRADI, M. & U. KRÜGER (1999): Können großflächige Beweidungssysteme als kostengünstige Naturschutzstrategie zur Erhaltung von bedrohten Offenlandarten dienen? Bericht über die Tagung "Großflächige Beweidungssysteme" am 8.10.1999 in Marburg. – Jahrbuch Naturschutz in Hessen 4: 189-195.
DÜLGE, R., ANDRETZKE, H. HANDKE, K., HELLBERND-TIEMANN, L. & RODE, M. (1994): Beurteilung nordwestdeutscher Feuchtgrünlandstandorte mit Hilfe von Laufkäfergesellschaften (Coleoptera: Carabidae). – Natur u. Landschaft 69(4): 148-156.
FIGURA, W., B. KRAHN, P. PAUSCHERT & B. SCHACKERS (1999): Pflege- und Entwicklungsplan für das Naturschutzgebiet „Körbecker Bruch“. – unveröff. Gutachten im Auftrage des Kreis Höxter – ULB
FISCHER, M., FUELLHAAS, U. & HUK, T. (1998): Laufkäferzönosen unterschiedlich anthropogen beeinflußter Feuchtgrünländer in vier Niedermooren Norddeutschlands. – Angewandte Carabidologie 1: 13-22.
FREUDE, H., HARDE, K. W. & LOHSE, G. A. (1976): Die Käfer Mitteleuropa. – Adephaga 1, 2, - Krefeld, Goecke und Evers, 301 S.
FRITZE, M.-A. (1998): Die Laufkäfergemeinschaften verschiedener Schilfröhrichte in Oberfranken (Bayern). – Angewandte Carabidologie 1: 83-94.
FUELLHAAS, U. (1998): Restitution von Feuchtgrünland auf Niedermoor - Der Einfluß mehrjähriger Überstau- und Vernässungsmaßnahmen auf Laufkäferzönosen. – Angewandte Carabidologie 1: 3-12.
HANDKE, K. (1995): Zur Laufkäferfauna eines Bremer Flußmarschengebietes (Niedervieland / Ochtumniederung / Ochtumsand). – Z. Ökologie u. Naturschutz 4: 203-225.
HEROLD, P. (2001): Auswirkungen einer Freiland-Schweinehaltung auf die Laufkäfer (Coleoptera, Carabidae) von Auengrünland. "Wenn der Bock zum Gärtner wird..." - Ergebnisse naturschutzorientierter Untersuchungen zum Thema Landschaftspflege durch Beweidung. – Wetzlar, NZH-Verlag: 225-283.
HEYDEMANN, B. (1957): Die Biotopstruktur als Raumwiderstand und Raumfülle für die Tierwelt. – Verh. Dt. Zool. Ges. Hamburg 20: 332-347.
HILL, B. T., BECK, L. A. & BEINLICH, B. (2003): Reaktionen der Laufkäferzönosen eines brachgefallenen Kalkackers auf extensive Schweinebeweidung. – Angewandte Carabidologie Supplement 3: im Druck.
HILL, B. T., BECK, L. A., BEINLICH, B. & DIETERICH, M. (2005): Veränderungen der Laufkäferzönosen (Coleoptera: Carabidae) verschiedener Grünlandtypen durch extensive Schweinebeweidung. in: Schweine in der Landschaftspflege –- Geschichte, Ökologie & Praxis. NEUGEBAUER, K.R., BEINLICH, B. & POSCHLOD, P. (Hrsg.), Stuttgart, Ulmer Verlag: im Druck.
HILL, B. T., BEINLICH, B., KÖSTERMEYER, H., DIETERICH, M. & NEUGEBAUER, K.R. (2004): The pig grazing project: Prospects of a novel management tool. in: Cultural Landscapes and Land Use. The Nature Conservation - Society Interface. DIETERICH, M. & VAN DER STRAATEN, J. (Hrsg.), DORDRECHT, Kluwer Academic Publishing: 193-208.
117
HILL, B. T., KÖSTERMEYER, H. & BEINLICH, B. (2000): Pigs – the forgotten grazing animal. – La Canada 13: 6-7.
IRMLER, U., SCHRAUTZER, J., GRABO, J., HANSSEN, U., HINGST, R. & PICHINOT, V. (1998): Der Einfluß von Nutzung und Bodenparametern auf die Biozönosen des Feuchtgrünlandes. – Zeitschrift f. Ökologie u. Naturschutz 7: 15-28.
KAMPF, H. (2000): Großherbivoren in großflächigen Beweidungssystemen - Erfahrungen aus den Niederlanden "Von der Politik zum Management". – Wageningen, Ministerium für Landwirtschaft, Naturschutz und Fischerei: 20.
KEGEL, B. (1990): Diurnal activity of Carabid beetles living on arable land. in: The role of ground beetles in ecological and environmental studies. STORK, N.E. (Hrsg.), Newcastle, Intercept: 65-76.
KREYER, D., HILL, B.T., KÖSTERMEYER, H., DIETERICH, M. & BEINLICH, B. (2000): Auswirkungen der Schweinefreilandhaltung auf ausgewählte Wirbellosenzönosen in verschiedenen Grünlandtypen – erste Ergebnisse. – Verh. Ges. Ökol. 30: 273.
KROKER, H. (1983): Beitrag zur Kenntnis der Bodenkäferfauna unbewaldeter Habitate der Warburger Börde (ohne Staphylinidae). – Abhandlungen aus dem Westfälischen Museum für Naturkunde 45(2): 3-15.
LARSSON, S.G. (1939): Entwicklungstypen und Entwicklungszeiten der dänischen Carabiden. – Entomol. Meddr. 20: 277-560.
LAUTERBACH, A.-W. (1964): Verbreitungs- und aktivitätsbestimmende Faktoren bei Carabiden in sauerländischen Wäldern. – Abh. Westfäl. Landesmuseum f. Naturkunde 26(4).
LEDERBOGEN, D.; G. ROSENTHAL, D. SCHOLLE, J. TRAUTNER, B. ZIMMERMANN & G. KAULE (2004): Allmendweiden in Südbayern: Naturschutz durch landwirtschaftliche Nutzung. – Angewandte Landschaftsökologie 62, BfN-Schriftenvertrieb im Landwirtschaftsverlag GmbH, Münster. 532 S.
LINDROTH, C.H. (1949): Die fennoskandischen Carabiden, eine tiergeographische Studie. III: Allgemeiner Teil. – Göteborgs Kungl. Vetensk. Vitterh. Sam. Handl. Ser., 4 (3), 911 S.
LOHR, M. (2005): Das NSG "Körbecker Bruch" bei Körbecke. – In diesem Heft.
LOMPE, A. (1989): Ergänzungen und Berichtigungen zu Freude-Harde-Lohse "Die Käfer Mitteleuropas" Band 2, U.O Adephaga. in: Die Käfer Mitteleuropas. LOHSE, G.A. & LUCHT, W. (Hrsg.), Krefeld, Goecke & Evers. 12: 23-59
LUFF, M.L. (1978): Diel activity patterns of some field Carabidae. – Ecological Entomology 3: 53-62.
MANDERBACH, R. (1998): Lebensstrategien und Verbreitung terrestrischer Arthropoden in schotterreichen Flußauen der Nordalpen – Marburg, Görich & Weihershäuser
MENKE, K. (2000): Die Laufkäferfauna von Röhrichten, Grünland und Grünlandbrachen im Werderland bei Bremen. – Angewandte Carabidologie 2/3: 19-36.
MICKLICH, D. (1996): Die Wirkung der Weidehaltung verschiedener Schweinerassen auf die Sukzession von Flußauengrünland und den physiologischen Zustand der Sauen. – Dissertation, Universität Rostock, Agrarwissenschaftliche Fakultät, 154 S.
MÜLLER, J., SCHMITZ, M., VOLPERS, M. & GERKEN, B. (1990): Beiträge zu einer Übersichtskartierung der Artengruppen der Springschrecken und Laufkäfer, als Grundlage für die Fortschreibung des Biotopkatasters und der Erstellung eines Schutzzielkonzeptes für den Kreis Höxter. – Höxter, Lehrgebiet Tierökologie, Gesamthochschule Paderborn.
NEUGEBAUER, K. (2003): Auswirkung der extensiven Freilandhaltung von Schweinen auf Gefäßpflanzen in Grünlandökosystemen. – Dissertation, Universität Regensburg, FB Biologie, 267 S.
RATHS, U. & U. RIECKEN (1999): Laufkäfer im Drachenfelser Ländchen. Tierwelt in der Zivilisationslandschaft - Teil III - Schriftenreihe für Landschaftspflege und Naturschutz, Bonn – Bad Godesberg, BfN, 156 S.
RIBERA, I., DOLEDEC, S., DOWNIE, I.S. & FOSTER, G.N. (2001): Effect of land disturbance and stress on species traits of ground beetle assemblages. – Ecology 82(4): 1112-1129.
RIECKEN, U. (1992): Planungsbezogene Bioindikation durch Tierarten und Tiergruppen - Grundlagen und Anwendung. - Schriftenreihe für Landschaftspflege und Naturschutz, 36, Bonn – Bad Godesberg, 187 S.
RIECKEN, U., FINCK, P. & SCHRÖDER, E. (2001): Tagungsbericht zum Workshop "Großflächige halboffene Weidesysteme als Alternative zu traditionellen Formen der Landschaftspflege". – Natur und Landschaft 76(3): 125-130.
118
|
RIECKEN, U., RIES, U. & SSYMANK, A. (1994): Rote Liste der gefährdeten Biotoptypen der Bundesrepublik Deutschland. – Schriftenreihe für Landschaftspflege und Naturschutz, Bonn, Kilda, 184 S. RUDOLPH, R. (1976): Coleoptera Westfalica: Familia Carabidae, Genera Leistus, Nebria, Notiophilus, Blethisa und Elaphrus. – Abhandlungen des Landesmuseums für Naturkunde in Münster / Westfalen 38(2): 3-22. SCHLEEF, J. & S. MIßFELDT (1998): Floristische und faunistische Kartierung im Gebiet „Rietbruch / Rösebecker Bruch“ 1997 (Kreis Höxter) – unveröff. Gutachten SCHÖLE, P. & H. TERLUTTER (1998): Rote Liste der in Nordrhein-Westfalen gefährdeten Sandlaufkäfer und Laufkäfer. – Angewandte Carabidologie 1: 51-62 SCHULZ, B. (2002): Die Reaktion ausgewählter Tierartengruppen (Coleoptera: Carabidae und Orthoptera: Acrididae) auf eine großflächige Extensivbeweidung am Beispiel der Weidelandschaft Oberes Eidertal. – Dissertation, Christian-Albrecht-Universität, Agrar- und Ernährungswissenschaftliche Fakultät. SMOLIS, M. (1982): Avifaunistische Bestandsaufnahme im geplanten Naturschutzgebiet "Körbecker Bruch". – Mitt. d. Naturkundl. Ver. Egge-Weser 1 (4): 142-182. THIELE, H.U. (1977): Carabid beetles in their environment. – Zoophysiology and Ecology, 10, Berlin, Springer, 369 S. TIETZE, F. (1973): Zur Ökologie, Soziologie und Phänologie der Laufkäfer (Coleoptera - Carabidae) des Grünlandes im Süden der DDR, II. Teil. Die diagnostisch wichtigen Carabidenarten des untersuchten Grünlandes und ihre Verbreitungsschwerpunkte. – Hercynia N.F. 10(2): 111-126. TRAUTNER, J. & GEIGENMÜLLER, K. (1987): Sandlaufkäfer – Laufkäfer. Illustrierter Schlüssel zu den Cicendiliden und Carabiden Europas. – Aichtal, Josef Margraf, 488 S. TRAUTNER, J., MÜLLER-MOTZFELD, G. & BRÄUNICKE, M. (1997): Rote Liste der Sandlaufkäfer und Laufkäfer Deutschlands (Coleoptera: Cicindelidae et Carabidae). – Naturschutz und Landschaftsplanung 29(9): 261-273. TURIN, H. (2000): De Nederlandse Loopkevers, Verspreiding en Oecologie (Coleoptera: Carabidae). – Nederlandse Fauna, 3, Leiden, Nationaal Naturhistorisch Museum Naturalis, 666 S. + CD-Rom. |
|
119